المدونة منشأة خصيصاً للرد علي شبهات الملاحدةو اللادينين و الربوبيين
و النصاري و اليهود غيرهم من الطاعنين في الاسلام وَاللَّهُ غَالِبٌ عَلَى أَمْرِهِ وَلَكِنَّ أَكْثَرَ النَّاسِ لَا يَعْلَمُونَ
يعتبر
السوط البكتيري بتركيبته المعقدة دليلاً كافياً على بطلان نظرية التطور .
فلو حصل وتعطل أو فقد جزء صغير من هذا النظام لعجز عن القيام بمهمته وأصبح
دون أدنى فائدة للبكتريا. يجب أن يكون عمل السوط متقناً منذ اللحظة التي
خلق فيها. هذه الحقيقة تلغي أيضاً المزاعم التطورية التي تعتمد مبدأ " التطور خطوة خطوة " . يدلنا هذا السوط على أن أشكال الحياة البدائية أيضاً تحمل تصميماً على جانب من التعقيد .
سأمثل فكرة السوط البكتيري بالمثال التالي
مثال الساعة التي تفتحها وتطلع على وظيفتها وما فيهاتروس بداخل الساعة وتراكبها وتكاملها مع بعضها البعض :فإننا سندرك - وبكل سهولة - معنى قولنا : ( التعقيد الغير قابل للاختزال ) ..وهو الذي يعني ذلك التعقيد والتركيب الذي لو أزلنا منه أي جزء : توقف الكل عن العمل !!..ودعونا ننظر مثلا ًلدائرة التروس البسيطة التالية :
والسؤال :
هل ترون أيا ًمن هذه التروس يمكن اختزاله - بمعنى الاستغناء عنه - :وتظل وتستمر نفس الحركة في الدوران ؟!!!..
فلينظر كل ٌمنا إليها جيدا ً: ويتخيل مثلا ًالاستغناء عن الترس الخارجي الأحمر الكبير ..
أو الأزرق ؟؟.. أو البنفسجي أو الأخضر أو الزيتي ؟؟؟..
نعم .. ففي كل الحالات سوف يتوقف التعقيد كله عن العمل إذا غاب أحد أجزائه !!!..
وهذا هو ما يجده الناظر في مكونات الساعة من الداخل !!..
وهذا بالضبط هو المقصود بتعبير :
(( التعقيد الغير قابل للاختزالIrreducible Complexity ))
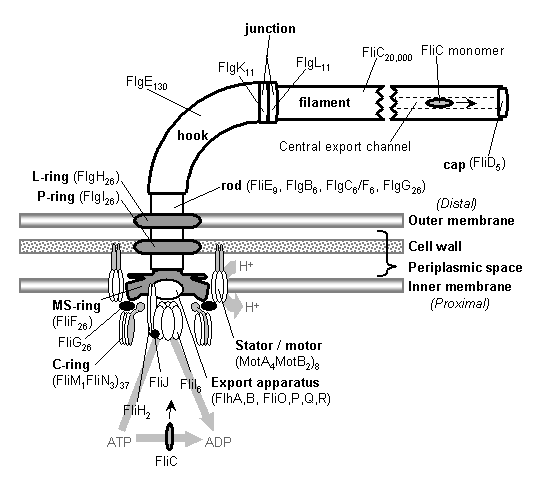
تستخدم
بعض أنواع البكتيريا ما يشبه السوط، ويطلق عليه " السوط " ليساعدها على
الحركة في المحيط السائل. يتصل هذا السوط بغشاء الخلية ويسمح للبكتريا حسب
الاتجاه الذي ترغب به بسرعة محددة .
عرف
العلماء السوط منذ زمن، إلا أن بنيته التي لم يكشف عنها النقاب قبل عقد
تقريباً، كانت مفاجأة كبيرة لهم . لقد اكتشفوا أن هذا السوط يتحرك من خلال "
محرك عضوي" في غاية التعقيد، وليس عن طريق آلية اهتزازية بسيطة كما كان
شائعاً.
تقوم بنية المحرك الدافع على مبدأ المحرك الكهربائي. هناك جزئين رئيسيين له : " ثابت " و " الدوران ".
يختلف السوط البكتيري عن باقي الأنظمة العضوية بأنه يولد حركة ميكانيكية . لا تستخدم الخلية الطاقة المختزلة في جزيئات ATP وإنما لديها مصدر خاص للطاقة : التي تستخدمها البكتيريا تنتج عن الأيوانات
المتدفقة عبر ِأغشية الخلية الخارجية . البنية الداخلية للمحرك معقدة جداً
. يشترك ما يقارب من 240 بروتين مختلف في بناء السوط . يتوضع كل منها في
مكانه المناسب بكل عناية . يعتقد العلماء أن هذه البروتينات هي التي تحمل
إشارات تشغيل المحرك وتوقيفه، وتشكل نقاط اتصال تسهل الحركة بمقاييس ذرية،
كما تفعل بروتينات أخرى مهمتها وصل السوط بغشاء الخلية. هذه الخصائص التي
يمتاز بها عمل هذا النظام تدل على الطبيعة المعقدة له
يعتبر
السوط البكتيري بتركيبته المعقدة دليلاً كافياً على بطلان نظرية التطور .
فلو حصل وتعطل أو فقد جزء صغير من هذا النظام لعجز عن القيام بمهمته وأصبح
دون أدنى فائدة للبكتريا .
يجب أن يكون عمل السوط متقناً منذ اللحظة التي خلق فيها. هذه الحقيقة تلغي أيضاً المزاعم التطورية التي تعتمد مبدأ " التطور خطوة خطوة " .
يدلنا هذا السوط على أن أشكال الحياة البدائية أيضاً تحمل تصميماً على جانب من التعقيد .
وكلما
تعمق الإنسان في دقائق هذا العالم، كلما اكتشف أن ما كان يظنه علماء
الأحياء في القرن التاسع عشر، بما فيهم داروين، أحياء أو عضيات بسيطة هي في
الواقع معقد تماماً كغيرها.
البروتينات المكونه للسوط
تصميم
معجز حتى في المخلوقات الصغيرة التي كان داروين وحاشيته يعتبرونها " بسيطة
" . يعتبر السوط البكتيري أحد الأمثلة التي لا تعد، تتحرك البكتريا في
الماء بتحريك هذا العضو المتصل بغشائها، كان الكشف عن التفاصيل الداخلية
لهذا العضو صدمة بالنسبة للعلماء تبين أن البكتيريا تحتوي عل محرك كهربائي
مقعد. هذا المحرك الكهربائي الذي يتكون من خمسة عشر جزيئاً مختلفاً هو
تصميم معجز في حد ذاته كما يظهر في الأعلى .
وكعادة التطوريون راحوا يرقعون بسيناريهوات كثيره لتفسير ذلك التعقيد الغير قابل للاختزال وكلها كانت محبطه حتى
ان البعض راح يشبهه الاهداب والزوائد ببعض الكائنات الدقيقه على انها
بدايه تطوريه لانتاج ذلك السوط ضاربا بذلك بداهات فى اى علاقه تطوريه
مفترضه سواء من ناحية التشابهه واحتمال التدرج التطورى او بالعلاقه
التطوريه بين الانواع وكيف يقتلها علم تقسيم تلك الكائنات وكان
اكثر السيناريوهات هو قولهم بان تلك الاجزاء قد تقوم بوظائف اخرى مثل
الحقن كما فى انواع كثيره من البكتريا وهو نظام يعرف ب بالنظام الإفرازي من
النمط الثالث او T3SS وهو نظام لافراز السموم وحقنها داخل الخليه المضيفه عن طريق ابرة ونظام حقن يشبهه التطوريون بالسوط الحركى فى
محاوله هزليه لاثبات قدرة تلك الاجزاء على التطور من وظيفه الى وظيفه اخرى
فيتحول الجهاز من الحقن المتخصص والسيطره على الخليه المضيفه الى سوط حركى
هيدروليكى فى سيناريو هزلى دون اعتبار ادنى سيناريوهات انتخابيه يمكنها ان
تستطيع ذلك او توجهه ولكن
الاشكال الاكثر غباءً والذى وقع فيه التطوريون بغض النظر عن تلك
التلكيكات هى انهم ايضا يواجهون نفس الاشكال التطورى مع جهاز الحقن المفترض
انه تطور للسوط لانه ايضا جهاز بالغ التعقيد وغير قابل للاختزال الوظيفى
التصميم المذهل في جهاز التعشيق (أو القابض) الجزيئي لسوط العصوية الرقيقة. كتبه
جوناتهان ماكلاتيشي:
Jonathan McLatichi
صاحب المقال: جوناثهان ماكلاتشي (Jonathan McLatchie) حاصل على ماجستير البحوث في البيولوجيا التطورية و النظامية في جامعة كلاسكوو Glasgow. حاليا هو محرر موقع evolution news""
لقد ظهر في الرسم التخطيطي بعض
الآليات الأساسية التي تتدخل في تركيبة السوط و في العملية الرائعة للجذب
الكيميائي البكتيري الذي بواسطته تغير البكتيريا اتجاهها استجابة للإشارات
الكيميائية.
هناك بكل تأكيد عدد كبير من
المتغيرات المختلفة في تلك النظم. إذا استنتجت أن هذه الآليات المدهشة
لديها تشابه كبير مع الأنظمة الذكية المستعملة في الهندسة فسوف يدهشك جهاز
التعشيق (أو القابض) الجزيئي لسوط العصوية الرقيقة. (Bacillus Subtilis) الموضح فوق من طرف بلير و آخرون (Blair) سنة 2008.
العصوية الرقيقة (Bacillus Subtilis) هي كائن حي قادر على تكوين بيوفيلم. البيوفيلم (Biofilm) هو عبارة عن مضادات حيوية مقاومة تفرزها البكتيريا مطعم في قالب من المواد البوليمرية خارج الخلية (EPS)
كالبروتينات و عديد السكريات الذي تلتصق فيه الميكروبات بعضها مع بعض. في
البيوفيلم تكون حركية البكتيريا غير مشغلة. و هو ما يجعلنا نطرح السؤال
حول كيف يتم منع هذه الحركية. كما يفسر ذلك الكاتب:
"السوط هو عبارة عن آلية جزيئية
معقدة، مقاومة و مكلفة و بكل بساطة فإن تغير تركيبة السوط لا يوقف بالضرورة
الحركية. بمجرد ما يتم تعطيل الجينات التي تحوي بنية السوط فذلك يستلزم
عدة جولات من الإنقسام الخلوي منذ أن يتم تركيب السوط الموجود من قبل إلى
أن ينقرض عند الخلايا الوليدة. في مقابل ذلك، فجهاز التعشيق (أو القابض)
يتطلب تركيب بروتين واحد لمنع الحركة. بالإضافة إلى ذلك إذا تم الإجهاض
المبكر لتكون البيوفيلم فإن السوط الذي كان متروكا يمكن إعادة تشغيله مع
تركيب المكونات السوطية. بينما أن التعبير الجيني و تركيبة السوط بطيئان و
معقدان فالتحكم في جهاز التعشيق (أو القابض) يعتبر أمر بسيط، سريع و قابل للإنعكاس بشكل كبير."
ما الذي يحدد إذا ما كان محرك السيارة متصلا أم لا بالأجزاء التي تمكن من التحكم في اتجاه العجلات؟ الجواب هو جهاز التعشيق (أو القابض) الذي يضمن بأن يكون المحرك و الترس غير متصلين. فسوط العصوي الرقيق، المشابه لجهاز التعشيق الموجود في السيارة هو عبارة عن بروتين يسمى EpsE، الذي من أجله يتدخل نفس المشغل الذي يراقب الجينات الضرورية لتكون EPS. فبروتين EpsE
يتواصل مع العضو الدوار للسوط (البنية التي تدور داخل العضو الثابت). و
البروتين الموجود في حالة البلمرة (أو الكوترة) داخل العضو الدوار يسمى FliG. فوحيدات FliG تحول الطاقة الصادرة عن تيار البروتونات في الطاقة الدورانية الخاصة بالسوط. عندما يتفاعل البروتين EpsE مع البروتين المبلمر FliG فذلك يولد تغيرا تكوينيا يسبب في انحراف يؤدي إلى انفصال العضو الدوار عن محرك البروتونات في السوط.
ف كوتينبلان و آخرون (Guttenplanl) 2010 أكد أن البروتين EpsE لديه وظيفتين إثنين: "يتفاعل مع العضو الدوار للسوط لمنع القابلية الحركية كما يتعاون مع إنزيمات أخرى لتركيب قالب EPS".
و لقد تمت الإشارة إلى ذلك في عدة مواضع و قد عبر البروفيسور دانييل كيمس (Daniel Keams) بيولوجي في جامعة بلومينكتون في إنديانا، و هو مدير البحث عن دهشته بالقول:
"الإكتشاف يمكن أن يعطي للمختصين لتقنيين في النانو أفكارا جديدة حول كيفية ضبط محركات صغيرة جدا في مجال الصناعة.
السوط هو واحد من المحركات الأصغر حجما و قوة في الطبيعة (...).يمكنه الدوران أكثر من 200 مرة في الثانية بفضل خاصية الدفع الدوراني بقدر 1400 بيكونيوتن-نانوميتر (Piconewton-nms). إنه كعدد كبير من أحصنة القوة لتشغيل آلة بقياس بضعة نانوميترات على مستوى العرض."
"إنه ليبدو لي شيء رائع على أن البكتيريا المتطورة و المهندسون البشر قد توصلوا إلى حل مماثل لنفس المشكل. كيف يمكن أن توقف بشكل مؤقت محركا قد بدأ بالإشتغال؟"
بكل تأكيد، من وجهة نظر التصميم الذكي، ليس من الغريب على أن يقوم جهاز تعشيق (أو القابض) بكتيري بمراقبة نظم متوازية مع ما تم تصميمه من طرف مهندسين بشر. ممكن أن يكون هناك تردد إذا ما تمت تسمية هذا النظام ب "التعقيد
الغير قابل للاختزال" (إنه شيء نسبيا بسيط و الأنزيم الوحيد الذي يشارك في
هذا النظام فهو يقوم بوظيفتين مختلفتين). إلا أنه لا يمكنك تجنب اعتبار
هذا النظام طرفا من التعقيد المحدد، كما يفسر ذلك دافيد طايلر في مدونته:
"فمقدار التعقيد يتجلى في التكوين المدهش للبروتين EpsE و قدرته في جذب البروتين الذي ينقل الدفع الدوراني مؤديا إلى عدم نقل هذه القوة. "
في نفس الوقت، فهذا النظام يحتاج
إلى العمل بتعاون مع آلية تفصل الحركة و تعيد الإرتباط بالمحرك. و كلما
تعلم المرء بشكل أكثر مبادئ الهندسة المتعلقة بالنظم البيولوجية، إلا و
كانت له دلائل أكثر تدفعه لاستنتاج أننا نتعامل مع أنظمة مصممة فعلا بعناية
فائقه.
محكمة كيتزميلر: السوط البكتيري، هل تم دحضه؟ الرد على الدليل الضعيف الذي قدمه كيتزميلر (الجزء الأول).
كاسي لاسكين (Casey Luskin)
التعريف بالكاتب:
كاسي لاسكين (Casey Luskin). محام، درس الماجيستير في العلوم و القانون. نال B.Sو M.Sفي
علوم الأرض بجامعة كاليفورنيا بسان دييغو. يعمل في معهد ديسكافري كمنسق
لمركز العلوم و الثقافة. كان قد قام من قبل ببحث جيولوجي في مؤسسة Scripps Institution for Oceanograpgy (1997-2002)
في قضية Kitzmiller v. Doverحكم القاضي جون إ جونس (John E. Jones)
بشكل قاس ضد الصحة العلمية للتصميم الذكي بإعلانه أن التعقيد الغير القابل
للاختزال لسوط البكتيريا الذي تم تقديمه من طرف المدافعين عن التصميم
الذكي خلال الحكم قد تم دحضه من طرف الشاهد و الخبير في البيولوجيا من
الجهة الداعية للدعوى الدكتور كينيد ميلر Kenneth Miller. هذا الأخير أول بشكل خاطئ
تعريف التعقيد الغير قابل للاختزال المشار إليه من طرف إحدى أشهر منظري
التصميم الذكي ميشيل بيهي عندما قدم حججه و فند حجج خصومه على أساس خاصية
ضعيفة جدا في البرهان. وفقا لما جرى، ميلر قام بطرح أن التعقيد الغير
القابل للاختزال يمكن دحضه إذا ما وجدت خارج ذاك النظام وظيفة أخرى مفصولة
عن نظام سابق لنظام معقد بشكل لا يقبل الاختزال، مقترحا بذلك أن النظام
السابق يمكن أن يكون قد تم إشراكه في النظام الأخير عبر العملية التطورية
في التكيف المسبق.
مع ذلك، فالتوصيف الذي قام به ميلر
يتجاهل تماما أن التعقيد الغير القابل للاختزال يتم تعريفه عن طريق إثبات
قدرة النظام الأخير على التطور في تسلسل عبر مراحل و هو ما لا يمكن إيجاد
أي وظيفة له في كل مرحلة من هذه المراحل. فقط يمكن تحديد ما إذا كان النظام
معقدا "تعقيدا يقبل الإختزال" أم "تعقيدا لا يقبل الاختزال" و ذاك بحذف
عناصر من النظام حتى يتم تحديد وظيفية كل مرحلة من المراحل الإنتقالية.
فالقدرة على البحث عن دور ما لجزأ معزول، كما هي وظيفة التطعيم التي يقوم
بها نظام الإفراز من النمط الثالث (الذي يتشكل تقريبا من ربع جينات السوط
البكتيري)، لا ينفي التعقيد الغير القابل للاختزال للنظام النهائي. علاوة
على ذلك، فميلر قد تجاهل أن التفسير التطوري لنظام ما يتطلب شيءا أكثر من
التكامل البسيط للأجزاء. و كتحليل أخير لما جرى، فإن شهادة ميلر لا تدحض في
الواقع التعقيد الغير القابل للاختزال حتى لا نترك المتتبعين لقضية
كيتزميلر (Kitzmiller) مع التصور الخاطئ على أن الأصل التطوري للسوط البكتيري قد تم حله.
تقديم: تعريف التعقيد الغير القابل للاختزال.
البيولوجي و منظر التصميم الذكي مايكل بيهي (Michael Behe)
يعرف التعقيد الغير القابل للاختزال مع بالأخذ في الإعتبار ما إذا كان
يمكن ملاحظة هل بإمكان نظام بيولوجي معين أم لا أن ينشأ إنطلاقا من عملية
تطورية عبر عدة مراحل. عرَّف بيهي التعقيد الغير القابل للاختزال في كتابه
"الصندوق الأسود لداروين":
"أعلن داروين في كتابه أصل الأنواع:
"إذا كان من الممكن إثبات وجود
أي عضو معقد لا يُرجَّح أن يكون قد تكَّون عن طريق تحولات عديدة ومتوالية
وطفيفة، فسوف تنهار نظريتي انهياراً كاملاً"
و النظام الذي يتفق مع معيار
داروين هو النظام المعقد تعقيدا لا يقبل الاختزال، و عندما أقول أنه معقد
بشكل لا يقبل الاختزال فأنا أقصد نظاما واحدا مكون من أجزاء مترابطة بشكل
صحيح تتفاعل فيما بينها من أجل تأدية وظيفة معينة بحيث إذا نقص أو تلف جزء
واحد فقط من هذه الأجزاء فكل النظام يتوقف عن العمل " (1)
خلال محاكمة Kitzmiller
فإن ميشيل بيهي شهد لصالح التصميم الذكي معطيا كدليل على ذلك أن السوط
البكتيري هو عبارة عن بنية بيولوجية تظهر فعلا تعقيدا لا يقبل الاختزال.
السوط البكتيري هو عبارة عن ليف يدور في شكل لولب مدفوع بمحرك يسمح
للبكتيريا بالتنقل في وسط سائل (2). و مع ذلك، عند النطق بالحكم فإن قرار
القاضي جونس اختلف مع تصريحات بيهي و زعم أن هذا الأخير يجهل كيف أن التطور
قادر فعلا على إنتاج بنيات معقدة كالسوط البكتيري:
"الدكتورين ميلر Miller و باديان Padian شهدوا أن مفهوم بيهي Beheبخصوص
التعقيد الغير القابل للاختزال يقوم على أساس جهل الأشكال التي يعمل بها
التطور. بالرغم من أن الدكتور بيهي أعطى تعريفا دقيقا للتعقيد الغير القابل
للاختزال عندما قال أن السابق الذي "ينقصه جزء واحد فقط فهو لا يصلح للعمل
أصلا" و ما يود أن يقوله هو بكل وضوح هو أنه سوف لن تكون له نفس الوظيفة
بالمقارنة مع النظام الذي يتوفر على كامل أجزائه. مثالا على ذلك، في حالة
السوط البكتيري فإن عزل جزء منه ربما سيؤدي إلى فقدانه لوظيفته كمحرك
دوراني. إلا أن الدكتور بيهي يستبعد في تعريفه إمكانية عمل سابق السوط
البكتيري ليس كمحرك دوراني ولكن بشكل آخر، كنظام إفراز" (3)
فيما يلي تقييم لنتائج القاضي بخصوص التعقيد الغير القابل للاختزال في سوط البكتيريا:
تجاهل التكيف المسبق؟
في قضية كيتزميلر، القاضي جونس استعمل مصطلح "التكيف المسبق" (Exaptation)
من أجل وصف كيف أن أي جزء يمكنه القيام بدور محدد في الخلية، فقط لكي يتم
استعماله فيما بعد من طرف نظام لا يقبل الإختزال من أجل القيام بوظيفة أخرى
مختلفة. في نص أكاديمي معروف جدا، "علم الأحياء التطوري" (Evolutionary Biology) يصف التكيف المسبق كالتالي:
"ستيفن جي كولد و إليزابيط فربا 1982 (Stephen Jay Gould and Elisabeth Vrba)
يشيران إلى أنه إذا كان إعتبار التكيف كميزة قد تطورت عن طريق الإنتخاب
الطبيعي و الذي يمتلك وظيفة عادية فذلك يتطلب مصطلح مختلف لتلك الخصائص
التي لم تتطور انطلاقا من تلك الوظيفة التي تقوم بها حاليا كما هو الحال
بالنسبة للعظام الجوفاء عند الطيور و خطوط إلتئام عظام الجمجمة عند
الثدييات اليافعة. هما يشيران إلى أن تلك الصفات التي تطورت إلى وظائف
أخرى، أو لكي لا تقوم بأي وظيفة على الإطلاق، و لكنهم اشتركوا من أجل
وظيفة جديدة فيجب تسميتهم ب"التكيف المسبق". " (4)
لقد ادعى القاضي في حكمه أن مايكل بيهي يجهل التكيف المسبق كوسيلة لتفسير أصل التعقيد البيولوجي:
"من خلال تعريف التعقيد الغير
القابل للاختزال بالشكل الذي قام به الأستاذ بيهي فهو يحاول إقصاء ظاهرة
التكيف المسبق انطلاقا من تعريفه متجاهلا بذلك الدلائل المتوفرة التي تدحض
برهانه." (5)
فقرار القاضي جونس على أن بيهي يجهل "التكيف المسبق" مبني على شهادة الدكتور كينيث ر.ميلر (Dr. Kenneth R. Miller)
التطوري و الشاهد الخبير في البيولوجيا و هو من الجهة المدعية خلال الحكم.
فالدكتور ميلر شهد أن التعقيد الغير القابل للاختزال سيتم دحضه إذا ما
استطاع أحد ما إيجاد أي وظيفة أخرى لكل جزء ثانوي للنظام بأكمله:
"افتراض بيهي مبني على أن أجزاء
نظام ذو تعقيد غير قابل للاختزال لا يمكن أن تكون لها وظائف مفيدة. و
بالتالي يجب أن نكون قادرين على أخذ السوط البكتيري و تفكيكه إلى أجزاء و
اكتشاف أن أيا من أجزائه لم تكن صالحة لأي شيء، إلا عندما نقوم بتجميعها
كلها في السوط" (6)
فتوصيف ميلر للتعقيد الغير للاختزال
غير دقيق على الإطلاق. خصوصا أن ميلر قد استعمل برهانه في حالات بيولوجية
واقعية حيث يؤكد أن بعض الأنظمة الثانوية للسوط البكتيري يمكنها أن تلعب
أدوارا أخرى في عدة كائنات حية. على سبل المثال، ميلر لاحظ أن نظام الإفراز
من الصنف الثالث (TTSS)
الذي يستعمل تقريبا 1/4 من الجينات المتداخلة في السوط (7) يمكن استعماله
من طرف بكتيريا مفترسة لحقن السموم في الخلايا حقيقيات النوى. وفقا لميلر،
فوجود (TTSS)
يظهر أن السوط البكتيري لا يعتبر نظاما لا يقبل الاختزال. مع ذلك، فتقديم
ميلر لبرهان نظام الإفراز من الصنف الثالث يحتوي ثلاث مشاكل رئيسية:
أ-الخبراء يؤكدون أن نظام TTSSهو من تطور من السوط و ليس العكس.
ب-بيهي
و غيره من أنصار التصميم الذكي قد كان لديهم علم مسبق منذ زمن طويل ب
"التكيف المسبق" و قد لاحظوا عدة مشاكل في هذا النوع من التفسيرات.
ت-ميلر صنف بشكل خاطئ كيف يمكن اختبار التعقيد الغير القابل للاختزال و لم يدحض سوى القش السطحي لهذا المفهوم المشار إليه من طرف بيهي.
1-من ظهر الأول؟ هل نظام TTSS أم السوط (أم أيا منهما)؟
أولا، يجب التذكير أن السلطة العلمية الرائد في النظاميات البكتيرية ميلتون ساير (Milton Saier) لا يزال يعتقد أن نظام الإفراز TTSS
هو من تطور من السوط و ليس العكس مما يجعل أن الطرح الذي قدمه ميلر يعتبر
مشكوكا فيه بشكل كبير. و بالرغم من أن ساير يعترف أن هناك من لا يتفق معه
إلا أنه لا يزال يؤكد أن TTSS قد تطور من السوط:
"بخصوص السوط البكتيري و TTSS فعلينا اعتبار ثلاث إمكانيات (و فقط ثلاثة): أولا: TTSS
ظهر أولا؛ ثانيا: نظام السوط هو من ظهر الأول؛ ثالثا: كلا النظامين تطوروا
من سابق مشترك. حاليا يوجد فقط القليل من المعلومات المتوفرة للاختيار من
الإمكانيات الثلاثة بدقة. و كما يحدث عادة عندما نقوم بفحص البراهين
التطورية فالباحث عليه أن يركز على الاستنتاج و الحدس المنطقيين. فحسب حدسي
الشخصي و البراهين التي نوقشت أعلاه فأنا أفضل الطريق الثاني. و أنت ما
رأيك؟" (8)
2-أجوبة واضحة لبيهي بخصوص إستئناف تفسيرات التكيف المسبق:
ثانيا، لدحض إدعاءات القاضي جونس
على أن بيهي "يحاول تجاهل ظاهرة التكيف المسبق إستنادا إلى تعريفه" و ما تم
عرضه من طرف ميلر على أن "افتراض بيهي يقوم على أن أجزاء نظام لا يقبل
الاختزال لا يجب أن تكون لهم وظيفة مفيدة،" فلنلق نظرة على بعض المقاطع من
كتاب: "العلبة السوداء لدارون" الذي يشير فيه بيهي إلى الصعوبات التي
تعتري البراهين الخاصة بالتكيف القبلي عندما يتحدث عن تطور الهدب:
"بما أن الهدب معقد تعقيدا لا
يقبل الاختزال فلا يوجد أي طريق مباشر يمكنه أن يؤدي إلى ظهوره. على هذا
الأساس، فالقصة التطورية للهدب يجب تصورها كطريق ثانوي أو غير مباشر، ربما
بتكيف الأجزاء التي كانت في الأصل مستعملة لغرض آخر." (9)
"مثالا على ذلك، افترض أنك تحاول
صناعة مصيدة فئران. ربما لديك في مرآبك قطعة خشبية كالتي يتم استعمالها
للامساك بالمثلجات عند بيعها (من أجل توفير القطعة الخشبية)، النابض عثرت
عليه في ساعة قديمة، و قطعة معدنية على شكل مُخْل (للمطرقة)، إبرة للخياطة
لاستعمالها كعارضة للإثبات و سدادة القنينة التي تحلو لك لاستعمالها كالفخ.
لكن هذه القطع لا يمكنها أن تشكيل مصيدة فئران صالحة من دون القيام
بتعديلات واسعة عليها، و حتى إذا ما حدث هذا التعديل فسوف لن تستطيع أن
تعمل كمصيدة للفئران. فالوظائف السابقة لهذه الأجزاء تجعلها غير مناسبة
للقيام بدور جديد كجزء من نظام معقد. في حالة الهدب توجد مشاكل مشابهة. فأي
بروتين متحول يتم ضمه إلى الأنابيب الدقيقة بشكل عشوائي سيؤدي إلى تعطيل
وظيفة هذه الأنابيب باعتبارها طرقا سيارة للتنقل. فالبروتين الذي يربط
الأنابيب الدقيقة فيما بينها بشكل عشوائي بإمكانه تشويه شكل الخلية – كما
أنه سيُشوَّه أي مبنى إذا ما تم وضع السلك بشكل خاطئ و الذي عشوائيا يمكنه
سحب الدعائم التي يرتكز عليها هذا المبنى. البروتين الرابط الذي يوحد بين
الأنابيب الدقيقة من أجل تكوين بنيات الإرتكاز فهو عادة يجعلها أقل مرونة
بخلاف النكسين الذي هو عبارة عن بروتين أكثر ليونة. فأي بروتين محرك غير
مضبوط جيدا إذا ما تسلل بشكل اعتباطي إلى هذه الأنابيب الدقيقة سوف يؤدي
إلى الضغط عليها و بالتالي إلى افتراقها عوض أن يجعلها تتحدب و تنغلق بشكل
ملائم. الهدب البدائي لا يمكن أن يكون قد ظهر على سطح الخلية أما إذا لم
يظهر هناك فإن النبض الداخلي يمكن أن يعطل الخلية؛ و لكن حتى إذا ما كان قد
ظهر على سطح الخلية فالبروتينات المحركة من المحتمل أنها كانت سوف لن تكون
كافية لتحريك الهدب. و حتى لوتحرك الهدب فأي حركة ضعيفة سوف لن تحرك
الخلية بالضرورة. و حتى لو تحركت الخلية فإن حركتها ستكون غير منظمة و سوف
لن تستجيب لأي حاجة من حاجيات الخلية." (10)
و قد أكد بيهي أيضا من قبل على أن التطور لا يتأتى دائما عبر الطريق المباشر:
"عند التحدث عن أي نظام ذو تعقيد
لا يقبل الاختزال (و كنتيجة لا يمكنه أن يكون قد تم إنتاجه بشكل مباشر)
فلا يمكن لأحد استبعاد إمكانية الطريق الغير المباشر لأن ذلك يزيد من تعقيد
النظام الذي يتفاعل فيه كل جزأ من الأجزاء المتداخلة في تركيبه و بالتالي
فإن إمكانية الطريق الغير المباشر تسقط بشكل سريع. و بما أن عدد الأنظمة
الغير القابلة للاختزال في تزايد فإن يقيننا بدحض معيار الفشل الذي وضعه
داروين يرتفع إلى أقصى ما يمكن أن يعترف به العلم." (11)
و كنتيجة لذلك، و على عكس تصريحات
القاضي جونس و ميلر، ترك بيهي على الطاولة إمكانية أن الأجزاء يمكن أن يكون
قد تم اختيارها بشكل مشترك انطلاقا من أنظمة أخرى و لا يبتعد كليا عن هذا
الإعتراض (فحتى الشريط الأساسي و التمهيدي بخصوص التصميم الذكي بعنوان "Unlocking The Mystery of Life"
فهو يتحدث عن برهان الخيار المشترك.). لقد شرح بيهي على أن وجود كل أجزاء
النظام لا يعتبر كافيا: فيجب التوفر على إرشادات التجميع المناسب لتلك
الأجزاء. و بالتالي فيجب التوضيح على أن ميلر قدم ميز بشكل خاطئ البراهين
التي قدمها بيهي متجاهلا دحض هذا الأخير لنظرية "الخيار المشترك" و مشيرا
بشكل غير صحيح أن بيهي قد قال "أن أجزاء النظام الغير القابل للاختزال لا
يمكن أن تكون لهم وظيفة مفيدة (خارج هذا النظام)".
3-التمييز الغير الصحيح لميلر بخصوص التعقيد الغير القابل للاختزال.
عند إعادة ما أكده ميلر يتبين لنا
أنه شهد على أن التعقيد الغير القابل للاختزال يمكن دحضه إذا ما استطاع
النظام الثانوي القيام بمهمة أخرى داخل الخلية:
"افتراض بيهي مبني على أن أجزاء
نظام ذو تعقيد غير قابل للاختزال لا يمكن أن تكون لها وظائف مفيدة. و
بالتالي يجب أن نكون قادرين على أخذ السوط البكتيري و تفكيكه إلى أجزاء و
اكتشاف أن أيا من أجزائه لم تكن صالحة لأي شيء، إلا عندما نقوم بتجميعها
كلها في السوط" (12)
فالسؤال يأخذ الشكل التالي: بماذا
يختلف الدليل الذي قدمه بيهي عن الذي قدمه ميلر؟ في الواقع فإن بيهي يطرح
التعقيد الغير القابل للاختزال كاختبار مع الأخذ في الاعتبار بنية النظام
بأكمله. فالتعقيد الغير القابل للاختزال يعمل على مجموعة من الأجزاء و ليس
على جزء واحد بمفرده. و حتى لو تم إيجاد وظيفة منفصلة للنظام الثانوي فهذا
الأخير لا يدحض النظام الغير القابل للاختزال و عدم القدرة على تطوير نظام
ككل.
فلنعد التعريف الذي قدمه بيهي:
"في كتابه "أصل الأنواع" ذكر دارون:
"إذا كان من الممكن إثبات وجود
أي عضو معقد لا يُرجَّح أن يكون قد تكَّون عن طريق تحولات عديدة ومتوالية
وطفيفة، فسوف تنهار نظريتي انهياراً كاملاً""
و النظام الذي يتفق مع معيار
داروين هو النظام المعقد تعقيدا لا يقبل الاختزال، و عندما أقول أنه معقد
بشكل لا يقبل الاختزال فأنا أقصد نظاما واحدا مكون من أجزاء مترابطة بشكل
صحيح تتفاعل فيما بينها من أجل تأدية وظيفة معينة بحيث إذا نقص أو تلف جزء
واحد فقط من هذه الأجزاء فكل النظام يتوقف عن العمل.
و على هذا المنوال، و وفقا لدارون،
فالتطور يحتاج لأن يكون النظام أو أجزاءه الثانوية قادرين على أداء وظيفة
ما خلال أي خطوة تطورية نحو تطور النظام النهائي حتى ولو كانت صغيرة. يمكن
لأحد ما أن يجد جزأ ثانوي وظيفي خارج النظام النهائي و حتى أنه يمكن لنظام
كلي أن يوجد مع عدة نقاط على طول "الطريق التطوري" و هو ما يطرح في الشك
فكرة القدرة على العمل عبر "تغيرات عديدة و متوالية و طفيفة" و لكنه من
الضروري تأكيد هذا الأمر حتى نستطيع في القريب التحدث عن التطور التدريجي.
بالتالي فإن ميلر ميز بشكل خاطئ
الدليل الذي قدمه بيهي كما لو أنه كان كل تركيزه منصبا حول لا وظيفية
الأجزاء الثانوية بينما أن البرهان الذي قدمه بيهي يركز بالفعل على قدرة
النظام بأكمله على التركيب بالرغم من أن الأجزاء المكونة له تمتلك وظائف
أخرى خارج وظيفة النظام النهائي.
ترقبوا ترجمة الجزء الثاني قريبا
التعريف بالكاتب:
كاسي لاسكين (Casey Luskin). محام، درس الماجيستير في العلوم و القانون. نال B.Sو M.Sفي
علوم الأرض بجامعة كاليفورنيا بسان دييغو. يعمل في معهد ديسكافري كمنسق
لمركز العلوم و الثقافة. كان قد قام من قبل ببحث جيولوجي في مؤسسة Scripps Institution for Oceanograpgy (1997-200
(8) Milton Saier, "Evolution of bacterial type III protein secretion
systems," Trends in Microbiology, Vol 12 (3) pg. 113-15, Marzo, 2004.
(9) Michael Behe, Darwin’s Black Box, pg. 65-66.
(10) Michael Behe, Darwin’s Black Box, pg. 66-67.
(11) Michael Behe, Darwin’s Black Box, pg. 40.
(12) شهادة الدكتور كينيت ميلر. اليوم 1. الفترة المسائية. ص.16
محكمة كيتزميلر: السوط البكتيري، هل تم دحضه؟ الرد على الدليل الضعيف الذي قدمه كين ميلر (الجزء الثاني)
السيارة كمثال توضيحي:
بهدف فهم كيف فشل اختبار ميلر في
تطبيق ما تمت صياغته من طرف بيهي بخصوص التعقيد الغير القابل للاختزال،
فلنأخذ مثال محرك السيارة و الترباس. فمحرك السيارة يحتوي على عدة أصناف من
الترابيس و كل ترباس يمكن اعتباره كجزء ثانوي صغير أو كنظام ثانوي في محرك
السيارة. فحسب منطق ميلر، إذا كان هناك أي ترباس أساسي في محرك السيارت
يقوم بوظيفة أخرى أيضا كالمقبض على سبيل المثال فهذا يدل على أن كل نظام
المحرك لا يعتبر نظاما معقدا بشكل لا يقبل الاختزال. فبرهان كهذا بكل وضوح
لا يمكن أن يكون سوى مجرد هراء.
لتحديد ما إذا كان المحرك نظاما
معقدا لا يقبل الاختزال فيجب التركيز على وظيفة المحرك نفسه و ليس على
أجزائه الثانوية التي يمكن أن تؤدي وظائف أخرى في مكان آخر. إنه من الواضح
أن الترباس خارج المحرك يمكنه القيام بدور آخر داخل السيارة إلا أن هذه
الملاحظة لا تفسر كيف أن هذه المكونات ذات التعقيد العالي كالمكابس و
الأسطوانات و عمود الكامات و الصمامات و عمود المرفق و شمعة الإحتراق و
غطاء الموزع و الأسلاك قد استطاعت أن تتصل فيما بينها وفق التعديل المناسب
بهدف صناعة محرك سيارة شغال. و حتى لو افترضنا أن هذه الأجزاء كلها قد تكون
لها وظائف أخرى داخل السيارة (و هو أمر مستبعد) فكيف استطاعت أن تتركب و
أن تتجمع بشكل مناسب لصناعة محرك سيارة شغال؟ فالجواب يستلزم تصميما ذكيا
بكل تأكيد.
بيهي قام بطرح أن النظام ذو
التعقيد الغير القابل للاختزال يتم تحديده إذا ما توقف عن العمل حين يتم
حذف جزأ من أجزائه. إنه اختبار مناسب لنظرية داروين لأنه يطرح السؤال
التالي: هل هناك مستوى أدنى مطلوب من التعقيد لكي يصير النظام وظيفيا؟ بكل
وضوح فمحرك السيارة يمتلك مجموعة من قطع الغيار الضرورية لاشتغاله. فقدرة
ترباس المحرك على أن يعمل كمقبض أيضا فذلك لا يفند التركيبة المعقدة الغير
القابلة للاختزال للأجزاء الضرورية لجعل النظام النهائي للمحرك شغال. فبيهي
لم يدعي أبدا أن الأنظمة الثانوية لا يمكنها القيام بوظيفة أخرى داخل
الخلية، فهو ادعى العكس من ذلك تماما. بل أكثر من ذلك فإن بيهي برهن بكل
بساطة أن التطور يدَّعِي أن النظام الإجمالي يجب أن يتشكل عبر مراحل طفيفة و
متوالية بحيث أن كل مرحلة تعتبر وظيفية.
قام ميلر بفهم التعقيد الغير
القابل للاختزال بشكل خاطئ و اختباره يعتبر ضعيفا حتى يستطيع دحضه. فاختبار
التعقيد الغير القابل للاختزال لا يسأل حول ما إذا كان بإمكان استعمال جزء
صغير من النظام الإجمالي من أجل القيام بوظيفة أخرى كما ادعى ميلر بل يطرح
السؤال التالي: هل يمكن للنظام بمجمله أن ينشأ عبر مراحل لا تتطلب فيها
تغيرات "غير طفيفة" للحصول على الوظيفة النهائية؟ فأي تعديل "غير طفيف" في
التعقيد يستلزم الإنطلاق من الأجزاء الثانوية الوظيفية التي تعمل خارج
النظام النهائي نحو النظام الإجمالي النهائي و الوظيفي الذي يمثل التعقيد
الغير القابل للاختزال.
و حتى لو استطاع ميلر إظهار أن
مكونا من مكونات السوط قد تواجد في أي مكان آخر في البكتيريا (و هو ما لا
يحدث في الواقع- فميلر لا يمكنه سوى التحدث عن الجسم القاعدي الذي يحوي
تقريبا 1/4 من بروتينات السوط) فميلر لم يعطي أي دليل معقول بخصوص تطور
السوط حتى يفسر كيف استطاعت مكوناته التجمع و التكون لتشكيل سوط وظيفي. و
حينها فقط يستطيع ميلر التأكيد على أن السوط البكتيري لا يعتبر معقدا بشكل
لا يقبل الاختزال.
.
خبراء آخرون يتفقون مع ميلر:
ويليام ديمبسكي William Dembski أدرك جوهر المشكل مع التعريف و التقييم الخاطئ لميلر بخصوص التعقيد الغير القابل للاختزال:
"إيجاد نظام ثانوي لنظام وظيفي
يقوم بوظيفة أخرى لا يعتبر دليلا مقنعا للبرهنة على أن النظام الأصلي قد
تطور من نظام آخر. فيمكن لأحد أن يقول بأن محرك الدراجة النارية يمكن
استعماله كخلاطة و بالتالي فإن محرك هذه الأخيرة قد تطور إلى محرك للدراجة
النارية. لعل ذلك ممكنا لكن ليس بدون تصميم ذكي. في الواقع، فإن الأنظمة
الوظيفية متعددة الأجزاء و المتداخلة بإحكام تحوي بشكل ثابت تقريبا أنظمة
فرعية متعددة الأجزاء و التي تقوم ببعض الأدوار الأخرى المختلفة. في أفضل
الحالات فإن نظام الإفراز من النمط الثالث TTSSيمثل
مرحلة ممكنة في التطور الغير المباشر للسوط البكتيري إلا أن هذا لا يعطي
حلا لمشكل هذا الأخير. فما نحتاج إليه هو مسار تطوري كامل و ليس فقط مجرد
واحة ممكنة على طول الطريق. فقول عكس ذلك سيكون كالذي يدعي إمكانية السفر
مشيا من لوس أنجلس إلى اليابان لأننا اكتشفنا جزر هاواي. فالبيولوجيا
التطورية تحتاج لفعل شيء أحسن من ذلك." (15)
بالرغم من أن ميلر قد شرح كيف يمكن أن يظهر بالكاد عنصر من العناصر المكونة للسوط فإن سكوت أ.مينيش و ستيفن سي ميير (Scott A.Minnich and Stephen C.Meyer) شرحوا أن مجرد توفر العناصر فذلك لا يعد كافيا لشرح تطور نظام بأكمله:
"حتى لو كانت العناصر
البروتينية متوفرة بشكل ما عند تشكيل محرك السوط خلال تطور الحياة،
فالعناصر بحاجة لأن تكون متشكلة وفق سلسة زمنية صحيحة كما تتشكل السيارة
داخل المصنع. حتى الآن و من أجل تجميع أجزاء محرك السوط فالبكتيريا الحالية
تحتاج لنظام مُحَضَّر من التعليمات الجينية بالإضافة إلى العديد من الآلات
البروتينية الأخرى للقيام بتنفيذ التعليمات الخاصة بالتجميع. منطقيا فأن
النظام نفسه يعتبر معقدا بشكل لا يقبل الاختزال. و على أي حال فإن برهان
التكيف المسبق يقتضي حاجة عدة أمور إلى تفسير- نظام بروتيني وظيفي بشكل
مستقل." (16)
على نفس المنوال، قام الفيلسوف أنجوس مينوج (Angus Menuge)
بتحديد عدد من العقبات التي يجب التغلب عليها عبر تفسيرات "التكيف المسبق"
و التي لم تتم الإشارة إليها من طرف ميلر خلال المحاكمة:
"لكي يظهر سوط بكتيري وظيفي عبر التكيف المسبق فقبل ذلك يجب مواجهة الشروط الخمسة التالية:
ش.1: الجاهزية. من بين العناصر
الجاهزة لتشكيل السوط فسوف نحتاج إلى التي لها القدرة على القيام بوظائف جد
متخصصة كالمجداف و الدوار و المحرك حتى و لو كانت لهذه العناصر وظائف أخرى
أم لا.
ش.2: التزامن. جاهزية هذه
العناصر يجب أن تكون متزامنة على هذا الشكل في نقطة ما بشكل انفرادي أم
مجتمعة، فكلها يجب أن تكون جاهزة في نفس التوقيت.
ش.3: التموقع: كل العناصر
المختارة يجب أن تتواجد في نفس "مكان البناء"، ربما ليس في نفس التوقيت و
لكن بكل تأكيد عندما يتم احتياجها.
ش.4: التنسيق. يجب أن تكون
العناصر متناسقة في الاتجاه الصحيح فقط: حتى ولو كانت كل عناصر السوط
متوفرة في الوقت الصحيح فإنه من الواضح أن معظم طرق التجميع ستكون غير
وظيفية أو دون أهمية.
ش.5: التوافق في الواجهة.
فالأجزاء يجب أن تكون متوافقة فيما بينها، أي "مركبة بشكل مناسب" و قادرة
على التفاعل بشكل صحيح: حتى و لو كان المجداف و الدوار و المحرك قد وضعوا
سويا في الترتيب الصحيح فهم أيضا سيحتاجون إلى امتلاك واجهة صحيحة. " (17)
ويليام دمبسكي(William Dembski) أخذ نهجا مماثلا لمينوج (Menugue).
فقام ديمبسكي فعلا بتجميع بعض معايير مينوج من أجل تطوير وسيلة لحساب
احتمالية تشكل شيء معقد لا يمكن اختزاله. فلحساب احتمالية وجود "شيء مميز و
مركب" فيجب الأخذ في الاعتبار احتمالية ظهور العناصر و احتمالية تواجدها
في نفس المكان بالإضافة إلى احتمالية تكون الأجزاء فيما بينها.
الجدول 1: مقارنة للمكونات التفسيرية المطلوبة لكل من مندومبسكي و مينوج ، مبنية على أي تكيف مسبق على حساب التطور.(18)
عامل ديمبسكي
الوصف
نظيره حسب معيار مينوج
احتمالية الظهور
احتمالية ظهور العناصر المشكلة للشيء.
الشرط الأول
احتمالية التموقع
احتمالية تموقع العناصر في المكان المطلوب عند ظهورها.
الشرط الثاني و الثالث
احتمالية التشكل
احتمالية تشكل العناصر المكونة للشيء فيما بينها عند ظهورها و تموقعها في المكان المناسب.
الشرط الرابع و الخامس
من الواضح أن ميلر قد وجد أن احتمالية ظهور 1/4 من بروتينات السوط قد يكون 1/1 إذا ما تواجد نظام الإفراز (TTSS)
(أو ربما بنية مشابهة) قبل السوط. إلا أنه لم يشر إلى أصل بقية البروتينات
الموجودة في السوط و لا حتى أشار إلى احتمالية التموقع أو إلى احتمالية
التشكل في السيناريو التطوري الذي قدمه. على ضوء معيار مينوج و ديمبسكي فمن
الضروري أن نطلب من ميلر أن يجيب على الأسئلة التالية:
-ما
هي الوظائف التي كان يقوم بها المجداف و الدوار و المحرك خارج السوط؟ (في
الحكم قام ميلر فقط بتفسير وظيفة الجسم القاعدي للسوط مشيرا إلى نظام
الإفراز من النمط الثالث TTSS و لكنه ترك الأجزاء المجهزة بمعدات الحركة الأكثر تعقيدا و حيوية في السوط دون تفسير.
-كيف تزامنت عناصر السوط؟ (خلال المحاكمة، ميلر لم يناقش هذا الأمر أبدا)
-كيف استطاعت هذه العناصر أن تتواجد في نفس "مكان البناء"؟ (أيضا لم يتطرق ميلر إلى هذا الأمر خلال المحاكمة)
-كيف
أن هذه العناصر استطاعت أن تتناسق بشكل بنيوي و أن تفاعل فيما بينها عندما
تجمعت للقيام بوظيفة السوط؟ (مرة أخرى ميلر لم يتطرق لهذا الموضوع)
خلال المحاكمة، ميلر لم يجب على
هذه الأسئلة المهمة. و بدلا من ذلك فقد قدم دليلا ضعيفا جدا بهدف دحض
التعقيد الغير القابل للاختزال و الذي عن طريقه أقنع القاضي لدرجة أنه لم
يقترب كليا من الإجابة عن كيف استطاع السوط أن يتطور فعلا.
القياس النهائي: القوس.
جواب ميلر بخصوص السوط البكتيري لا
يدحض تعقيده الغير القابل للاختزال لأن ميلر لم يشر إلى أي شيء بخصوص كيف
استطاع النظام السوطي النهائي الظهور. حتى لو وجدت وظائف أخرى لنظام
الإفراز من النمط الثالث TTSS
هذا لا يعني أن النظام السوطي سوف لن يحتاج لقفزات كبيرة في التعقيد (أو
باستعمال مصطلحات داروين: تغيرات غير طفيفة) من أجل إنتاج سوط وظيفي في آخر
المطاف. و لاستخدام القياس النهائي لإظهار عدم صحة تفسيرات ميلر فلنأخذ
مثالا لمحاولة القيام ببناء قوس معقد بشكل لا يقبل الإختزال:
الصورة "أ": هذا السوط معقد بشكل لا يمكن اختزاله، فإذا ما تم حذف قطعة
واحدة فقط منه فإن البقية سوف تسقط
(ملاحظة: لغرض التوضيح، فأنا أجهل
مؤقتا مسألة أن قوسا معقدا لا يقبل الاختزال يمكن أن يكون نتاجا لعمليات
التعرية الطبيعية. فأنا أعي جيدا أن هذا القياس لا يناسب كثيرا حقل
البيولوجيا و لكن ليس الهدف من هذا التوضيح دحض تلك المسألة.)
وفقا لميلر، إذا ما استطعنا إيجاد
وظيفة أخرى لعنصر ثانوي فذلك يعني أن نظام لا يعتبر معقدا بشكل لا يمكن
اختزاله. حسنا، فلنقم الآن بتفكيك هذا القوس إلى قطع ثانوية.
الصورة
"ب": هنا قمنا بتقسيم السوط إلى أجزاء ثانوية. بشكل مشابه، فقد وجد ميلر
ظاهريا جزءا ثانويا في السوط "نظام الإفراز من النمط الثالث" و الذي
بإمكانه القيام بوظيفة أخرى و نظام الإفراز من النمط الثالث لا يحوي أكثر
من 1/4 من مجموع العناصر السوطية. بشكل مماثل، في هذا القوس توجد قطعة
ثانوية سميت بحرف "س" و التي تشكل 1/4 من مجموع القوس و التي بإمكانها أن
تقوم بوظيفة أخرى خارج القوس مثلا يمكنها البقاء واقفة هنا بنفسها. إلا أن
هذا يظهر عدم صحة اختبار ميلر، فقدرة القطعة الثانوية "س" هذه على الوقوف
لوحدها لا يعني ذلك على أن القوس لا يعتبر معقدا بشكل لا يقبل الإختزال و
بالتالي فإذا حذفنا القطعة الثانوية "ت" الموجودة في الأعلى فإن القوس
سينهار حتى و لو بقيت القطعة "س" واقفة.
الصورة
"ت": حتى ولو امتلكت القطعة الثانوية "س" وظيفة أخرى خارج نطاق القوس كأن
تبقى واقفة لوحدها فذلك لا يعني أن القوس لا يعتبر معقدا بشكل لا يقبل
الاختزال بحيث يمكن أن يتشكل عبر عدة مراحل. و بالتالي فإن الاختبار
المناسب للتعقيد الغير القابل للاختزال يكون بالسؤال حول ما إذا كان النظام
بأكمله يمكن أن ينشأ مرحلة بعد أخرى من خلال تغيرات طفيفة و خفيفة. فمن
المهم إدراك أن النظام لا يتحول إلى "تعقيد قابل للاختزال" فقط لأن جزءا من
أجزاءه يبقى وظيفيا خارج النظام النهائي.
الاختبار الصحيح للتعقيد الغير القابل للاختزال:
اختبار ميلر هو اختبار بسيط و سطحي و شخصي، و حتى يكون صحيحا فكان يجب صياغته على الشكل التالي:
"إن تنبؤ بيهي يرتكز على أن
النظام المعقد بشكل لا يقبل الاختزال سيسير عبر عدة مراحل غير وظيفية على
طول أي مسار تطوري. و بالتالي فعلينا أن نكون قادرين على أخذ السوط
البكتيري على سبيل المثال و حذف جزء منه و اكتشاف أن النظام يتوقف عن
العمل."
إن شهادة ميلر و استنتاج القاضي جونس (Jones)
تم بناءهما على اختبار خاطئ للتعقيد الغير القابل للاختزال و الذي يركز
على وظيفية الجزء الثانوي و ليس على وظيفية النظام السوطي بأكمله. إذا ما
استطاع ميلر إيجاد وظائف موازية لكل النظم الثانوية للسوط سيكون قد أحرز
على الأقل تقدما لتفسير تطوره لأنه بذلك سيكون قد استجاب للمعيار الأول
لمينوج (Menuge) المتعلق ب "الجاهزية" (الشرط الأول). لكن و كما تسألا مينيش (Minnich) و ميير (Meyer):
"و البروتينات الثلاثون الأخرى الموجودة في المحرك السوطي (و التي لا توجد في نظام الإفراز من النوع الثالث TTSS) هي خاصة بالمحرك فقط و لا توجد في أنظمة كائنات أخرى. إذن: من أين تكيفت بشكل مسبق هذه العناصر البروتينية؟"
ميلر لم يجب على هذا السؤال. و كما تمت الإشارة، فكل ما استطاع ميلر أن يقدمه هو فقط TTSS كدليل على تطور السوط و أشار فقط إلى جاهزية 1/4 من المعيار الأول لمينوج متجاهلا إمكانية احتمال تطور TTSS
من السوط و ليس العكس. و إذا ما استطاع ميلر الجواب على "جاهزية" (ش.1)
كل عناصر السوط ، فسوف يجب عليه بعدها إعطاء تفسير للتزامن (ش.2) و للتموقع
(ش.3) و للتنسيق (ش.4) و للتوافق (ش.5) حتى يجيب عن مسألة تطور السوط و
هكذا القضاء على الحاجة إلى تعديلات "غير طفيفية" على طول مساره التطوري.
لكن ميلر لم يقم بأي شيء من هذا. و
بالرغم من عدم كفاية تفسيراته فإن القاضي جونس اعتبر أن أدلته و تصريحاته
الغير الدقيقة بخصوص التعقيد الغير القابل للاختزال كانت صحيحة. لقد تقرر
هذا مع الضرب بعرض الحائط للشهادة الموسعة التي قدمها خلال المحاكمة عالم
الأحياء الدقيقة و المساند للتصميم الذكي و الخبير في السوط الدكتور سكوت
مينيتش (Dr.Scott Minnich)
و قد أدلى بها لإثبات التعقيد الطبيعي الغير القابل للاختزال للسوط. هذه
شهادة مينيتش التي تم تجاهلها بشكل كامل من طرف القاضي خلال قضية كيتزميلر:
"أنا أعمل على السوط البكتيري
لفهم وظيفته، على سبيل المثال، من خلال تعريض الخلايا على مركبات مطفرة أو
على عدة وسائط لتسجيل تلك الخلايا التي ضعفت أو فقدت حركيتها. هذا هو نمطها
الظاهري، فالخلايا تستطيع أو لا تستطيع السباحة. فنحن نقوم بتعريض الخلايا
للطفرات: فإذا أصبنا جينا له علاقة بوظيفية السوط فإن الخلايا سوف لن
تستطيع السباحة و هذا هو النمط الظاهري القابل للتسجيل الذي نستعمله. ثم
تستعمل بعد ذلك الهندسة العكسية لتحديد كل هذه الجينات و نربط هذا
بالكيمياء الحيوية لإعادة تركيب البنية بالأساس و لفهم ما هي وظيفة كل جزء
على حدة. باختصار فإن هذه العملية هي مشابهة للتصميم الذي دفع البيولوجيا
من علم وصفي بسيط إلى علم تجريبي من حيث توظيف هذه التقنيات." (20)
مينيتش شرح أنه عرض كل جينات السوط
للطفرات و وجد أن السوط يفقد وظيفيته إذا ما فقد جين واحد فقط. و بالتالي
فإن السوط يمتلك تعقيدا لا يقبل الاختزال في ما يخص تكامل الجينات:
"طفرة واحدة تقضي على جزء واحد
فلا تستطيع الخلية السباحة. فقمت بوضع هذا الجين مرة أخرى في مكانه فتعود
الحركية من جديد. أزلت جزءا و وضعت مكانه مرة أخرى نسخة جيدة من الجين و من
جديد فهي تستطيع السباحة. فتعريف هذا هو أن هذا النظام معقد بشكل لا يقبل
الاختزال. لقد قمنا بهذا الأمر مع كل العناصر 35 المكونة للسوط و دائما
يعطينا نفس النتيجة." (21)
و لكن للأسف فإن القاضي جونس قد قرر جهل هذه الشهادة.
الخاتمة:
مع الأخذ في الاعتبار قرارات
القاضي جونس فإن الأدلة الموالية للتصميم الذكي المتعلقة بالتعقيد الغير
القابل للاختزال الموجود في السوط لم يتم أبدا دحضها كما ادعى القاضي.
فميلر قدم للقاضي تصوير خاطئ بخصوص التعقيد الغير القابل للاختزال و منهاج
سطحي جدا لاختباره. للأسف فإن قرار القاضي الذي كرس تحريف ميلر سيترك
المجتمع العلمي و الناس بشكل عام مع فكرة أن الأصل التطوري للسوط البكتيري
قد يمكنه تفسيره. و بشكل لا يصدق و بالرغم من شهادة مينيتش و تقديمه
لمنجزاته التجريبية فإن القاضي جونس ادعى أن "التصميم الذكي...فشل في...الاشتراك في البحث و في الإختبار." (22)
فالقاضي تقبل أيضا ادعاء ميلر
الخاطئ على أن التعقيد الغير القابل للاختزال لا يعتبر برهانا إيجابيا لدعم
التصميم و لكن فقط برهانا بسيطا و سلبيا ضد التطور. ربما نمتلك نظرات
مختلفة بهذا الخصوص إلا أن مينيتش و ميير قد جعلا منه مثالا إيجابيا
للتصميم في السوط:
"تظهر الآلات الجزيئية توقيع
أصلي أو سمة مميزة للتصميم تدعى بالتعقيد الغير القابل للاختزال. في جميع
الأنظمة ذوات التعقيد الغير القابل للاختزال و التي تعرف فيها وظائف النظام
من خلال التجربة أو الملاحظة فإن الهندسة أو التصميم الذكي قد لعبوا دورا
أساسيا في ظهور النظام. فكوننا وجدنا أنظمة بيولوجية تقلد قدراتنا في
التصميم و الهندسة فذلك يعطينا تفسيرا للسؤال حول ما إذا كانت هذه النظم هي
نتاج للحظ و للحاجة الغير الموجهة من دون أي هدف. في الواقع و في أي سياق
آخر سوف نعترف على الفور بأن تلك النظم هي نتاج لهندسة ذكية. بالرغم من أن
البعض سيقول بأن هذا دليل بسيط مبني على الجهل، إلا أننا نعتبر ذلك
كاستنتاج لأفضل تفسير بما أننا نعرف أن الظواهر الذكية هي مناقضة تماما
للأسباب الطبيعية أو المادية." (23)
و في الأخير فإن قرار القاضي جونس
حول أصل السوط البكتيري يجب تجاهله لأنه مثال للسياسة الحزبية و ليس
للتحليل العلمي الدقيق و الموضوعي.
أخص بالشكر أليكس بينز (Alex Binz) و دافيد كلينغوفر (David Linghoffer) و كيميائي حيوي مجهول من جامعة كاليفورنيا لمساعدتهم لي في إنجاز هذا المقال. كل الأخطاء الموجودة تنتسب لي.
لمذا لم يفند البرهان الذي قدمه بيهي بخصوص السوط البكتيري؟
جوناتهان ماكلاشي. Jonathan McLachie
إن الذين كرسوا بعضا من وقتهم
لقراءة الأدبيات الخاصة بالجدال القائم حول التصميم الذكي و التطور سيكونون
معتادين بكل تأكيد على الجواب الذي قدمه التطوريون فيما يخص برهان التعقيد
الغير القابل للاختزال الذي قدمه بيهي و الذي يتحدث فيه عن السوط
البكتيري. على ما يبدو، هناك إجماع في الرأي بين أغلب التطوريين على أن
التعقيد الغير القابل للاختزال الموجود في السوط البكتيري قد تم دحضه و على
أننا نحن أنصار التصميم الذكي نسعى لتغيير قواعد اللعب بشكل مستمر و ذلك
بدفن رؤوسنا في الرمال و بتشبتنا بأدلة جد سطحية. و كما كان متوقعا، قام
أحدهم مؤخرا بكتابة ما يلي عبر موقع الفيسبوك:
"ما يزعجني أكثر في أنصار التطور
هو عدم استسلامهم أبدا. كم مرة يجب علي أن أقول لك على أن هذا الأمر غير
صحيح حتى تتقبله؟ كم مرة تم دحض التصميم الذكي عبر أبحاث معترف بها حتى
تعترف على أنها قضية خاسرة؟ إن تاريخ التعقيد الغير القابل للاختزال
الموجود في السوط البكتيري يعتبر بشكل مطلق و نهائي أمرا مغلقا. إن ذلك لم
يكن سوى مهزلة. و إن اللعبة قد انتهت."
مؤخرا عندما كنت في أحد المطاعم
الأدبية التي كانت تحتضن جلسة حول الإيمان و العلم قمت بتقديم السوط كمثال
للتعقيد الغير القابل للاختزال و تلقيت أجوبة مماثلة.
لكن، هل يعتبر هذا الإدعاء صحيحا
فعلا؟ هل تم دحض هذا الدليل؟ منذ حوالي عام قرأت كتابا بعنوان: "لماذا فشل
التصميم الذكي. نقد علمي للخلقيين الجدد." (Why Intelligent Design Fails- A Scientific Critique of the New Creacionism) كتبه كل من مات يونغ و تينر إيديس (Matt Young and Taner Edis) و قد ساهم في كتابة الفصل الخامس من هذا الكتاب يان موسغريف (Ian Musgrave) و الذي يحمل عنوان: "تطور السوط البكتيري" و الذي يحاول فيه الرد على ميشيل بيهي و ويليام دومبسكي (Michael Behe and William Dembski)
بشكل قطعي و حاسم فيما يخص التعقيد الغير القابل للاختزال. فبينما كنت
أقرأ هذا الفصل أتذكر عدم شعوري بأي انطباع. ففي الصفحة 82 يقدم موسغريف
البرهان التالي:
"هذا هو السيناريو المحتمل لتطور السوط البكتيري: يظهر أولا نظام الإفراز قائم على قضيب SMC
و على تركيب معقد من المسام، و الذي يعتبر الأصل المشترك بين نظام الإفراز
من النمط الثالث و النظام السوطي. و بإشراك مضخة أيونية (و التي ستتحول
فيما بعد إلى بروتين للمحرك) إلى هذه البنية سمحت بتطوير وظيفة الإفراز.
حتى اليوم، فبروتينات المحرك و التي تنتمي لعائلة من البروتينات تقوم
بالإفراز يمكنها الإنفصال و الاتحاد من جديد مع بنية السوط بكل سهولة.
فالقضيب و المسام ذو التركيب المعقد ربما كان بإمكانهم الدوران في هذه
الحال كما يقع في بعض الأنظمة حيث تكون الحركية عن طريق الانزلاق دون
امتلاك محرك. لقد ظهر ليف السوط البدائي فيما بعد كجزء من بنية الإفراز
البروتيني (مقارنة مع الشعرة الزائفة (و هي جزء من البروتيوبكتيريا غاما) و
الدودات الزائدة ذوات الألياف الموجودة في السلمونيلا و بنيات الألياف
الموجودة في الإشريكية القولونية). فالحركية المتناوبة بين الانزلاق و قوة
الدفع قد ظهرت خلال المرحلة التالية أو التي بعدها لتستقر بعدها عند حركة
السباحة. يمكن أن يضاف التنظيم و التحول فيما بعد لأن هناك بكتيريا معاصرة و
التي بالرغم من عدم توفرها على هذه الصفات فهي تعمل بشكل جيد في بيئتها
(شاه و ساكت 1995/ Shah and Sockett 1995). في كل مرحلة هناك فائدة للتغيرات في البنية."
في الواقع، فإن مارك بالن و نيك ماتزك (Mark Pallen and Nick Matzke) أدلوا بحجة مشابهة في مقال لهم بمجلة الطبيعة "Nature"
عام 2006 (و قد تمت الإشارة إليه من طرف إحدى الجمهور المستمع خلال
الزيارة الأخيرة لبيهي إلى المملكة المتحدة). و يُعرَف كين ميلر أيضا
بادعاءاته الروتينية المشابهة بخصوص السوط البكتيري انطلاقا من نظام
الإفراز من النوع الثالث و الذي يقوم أساسا على اعتبارات التشابه في
التسلسلات البروتينات.
إذن، هل بإمكان هذه النقط أن تدحض
بشكل نهائي العمل المزعج و المقلق الذي يطرحه التصميم الذكي؟ فهي لا
تستطيع. لأن حجج كل من السادة المذكورين آنفا يستهينون بعدة مسائل مهمة.
أولا و قبل كل شيء، فهم يستهينون
بالتعقيد و بالتركيبة الهائلة للنظام السوطي – كجهاز التجميع و التصميم
الفني لبنيتها-. في الواقع، فإن العملية التي يتشكل عن طريقها السوط بشكل
ذاتي داخل الخلية تعتبر جد معقدة بحيث يجب علي القيام بمجهود كبير حتى
أشرحه بشكل مفهوم للأشخاص الذين لا يعلمون شيء بخصوصه. فهذه المفاهيم
الأساسية تعتبر صعبة للاستيعاب من طرف أولئك الذين ليسوا معتادين على تصور
نظام كهذا أو من طرف أولئك الذين يدرسونه لأول مرة. لكن، و في نفس الوقت،
فالأساسات الميكانيكية لتجميع السوط تعمل برشاقة و سحر رائعين كالهندسة
الرائعة الموجودة في المحرك السوطي –و هو التحدي الكبير الذي يطرح للتطور- و
التي لا يمكن تقييمها بشكل صحيح دون حد أدنى من المعرفة السطحية لعملياتها
المدعمة. فلنلق نظرة موجزة.
التجميع الذاتي للجهاز السوطي:
تركيبة السوط البكتيري تتطلب
تعبيرا منظما لأكثر من 60 منتوجا جينيا. فتركيبته الحيوية داخل الخلية تنظم
عن طريق جينات مرتبة في تسلسل منظم بشكل دقيق و التي فيها التعبير عن جين
واحد في مستوى معين يستلزم التعبير المسبق لجين آخر في مستوى عال. و كنموذج
للتجميع السوطي نجد السالمونيلا و هي صنف من البكتيريا المعوية و بالتالي
فإن مناقشتي سترتكز على هذه البكتيريا ما لم يرد خلاف ذلك.
النظام السوطي للسالمونيلا يتوفر
على ثلاث أصناف من المحفزات (المحفز يشبه نوعا من المفتاح المفصلي الجزيئي و
الذي بإمكانه بدأ التعبير الجيني عندما يتم التعرف عليه من طرف الحمض
الريبوزي النووي ARNو
من طرف بروتين مختص متداخل معه يسمى "عامل سيغما"). و هذه المحفزات تعرف ب
"الصنف 1" و "الصنف 2" و "الصنف 3" و نسخها التسلسلي مرتبط بعملية التركيب
السوطي. فالصنف 1 يحوي جينين اثنين في مشغل واحد (يسميانFlhD و FlhC).
و الصنف 2 يحوي 35 جين موزع على ثمانية مشغلات (بما في ذلك الجينات
المسؤولة عن تجميع الخطاف بالجسم القاعدي و بعناصر سوطية أخرى بالإضافة إلى
جهاز التصدير و جينين اثنين آخرين منظِّمين يسميان "FliA" و "FlgM"). أما الجينات الأخرى المسؤولة عن تشكل الليف فيتم التحكم فيها من خلال محفزات الصنف الثالث.
محفز الصنف 1 يسوق التعبير عن المنظم الرئيسي (خاص بالبكتيريا المعوية التي تنتمي إليها السالمونيلا) المسمى "FldH4C2" (لا تقلق بشأن تذكره) و حينما يبدأ بالعمل فالمنظم الرئيسي يعمل على محفزات الصنف 2 و يقوم بذلك بالاشتراك مع "عامل سيغما" σ70 (للتذكير فلقد أشرت من قبل أن "عامل سيغما" ل ARN
هو عبارة عن بروتين قادر على الترابط بشكل محدد مع المحفزات الجينية).
فمحفزات الصنف 2 هي المسؤولة عن التعبير الجيني للوحدات الفرعية و للمنظمين
للخطاف و للجسم القاعدي بالإضافة إلى عامل سيغما آخر يسمى σ28(و الذي يشفر من قبل جين يسمى FliA) و عامل سيغما مضاد FlgM (كما يشير إليه اسمه فعامل سيغما مضاد يتحد مع عامل سيغما بهدف منع نشاط الاستنساخ). أما عامل σ28
فهو ضروري لتنشيط محفزات الصنف 3 و هنا بالضبط أين نجد أنفسنا في مشكل إذ
لا يوجد أي معنى للبدء بتركيب مونومرات الفلاجلين قبل أن يتم تركيب الخطاف و
الجسم القاعدي و بالتالي و بهدف كبح σ28فعامل سيغما المضاد (FlgM) و الذي تمت الإشارة إليه سابقا فهو يكبح نشاطه و يمنعه و ذلك بالعمل على التفاعل الذي يتشكل مع الهولوأنزيم الموجود في ARN. و عندما يتم الإنتهاء من تشكل الخطاف و الجسم القاعدي فعامل سيغما المضاد FlgM
يتم إفرازه عبر البنيات السوطية التي تم إنتاجها من طرف جينات الصنف 2، و
بعد ذلك و أخيرا فإن محفزات الصنف 3 (المسؤولة عن تعبير مونومرات الفلاجلين
و عن الانجذاب الكيميائي و المحركات المولدة للقوة) يتم تشغيلها من طرف σ28و يمكن الانتهاء من السوط.
لكن هذا يصبح أحسن بكثير، فنظام التصدير السوطي (الذي عن طريقه يتمحذف FlgM من الخلية) لديه حالتين من نوعية الركائز (Substrate):
ركائز من نوع القضيب/ الخطاف و ركائز من نوع الليف. فخلال عملية التجميع
السوطي، فإن "مفتاح" نوعية الركيزة يجب أن يكون مدفوعا حتى يتغير من الحالة
السابقة إلى النهائية. و البروتينات التي تشكل أجزاء من الخطاف و من
القضيب يجب أن يتم تصديرها قبل تلك التي تشكل الليف. لكن، كيف يأخذ مكانه
هذا التغير في نوعية الركيزة؟
و العنصر الرئيسي في هذه العملية هو بروتين مرتبط بالغشاء يسمى FlhB. كما يوجد كذلك بروتين منظِّم لطول الخطاف يدعى FliK و الذي يقوم بالتأكد من أن طول الخطاف يوجد في القياس الصحيح (55 nm) و نفس البروتين مسؤول أيضا عن تغير نوعية ركيزة التصدير. و كما اتضح، فمن دون Flik ستفقد القدرة كليا على تغيير و تصدير الليف و على مراقبة طور الخطاف. فبروتين Flik لديه مجالين أساسيين: مجال N-terminal و مجال C-terminal فخلال تجميع الخطاف ف FliKN يعمل كجهاز استشعار جزيئي و يرسل معلومات حول طول الخطاف و عندما يَبلُغ هذا الأخير الطول الصحيح تنقل المعلومة ل FliKC و FliKCTو هو ما يفضي إلى تغير تركيبي و هذا بدوره يؤدي إلى ارتباط FliKCT ب FlhBC، و هذا الترابط يفضي إلى تغير تركيبي في FlhBC. و هذا ما يسبب تغير في نوعية الركيزة.
فالتجميع السوطي يبدأ في الغشاء
السيتوبلازمي ثم يتقدم عبر الحيز المحيط بالغشاء الهيولي ليمتد أخيرا خارج
الخلية. فالسوط يتشكل أساسا من جزأين أساسيين اثنين: نظام الإفراز و البنية
المحورية. و المكونات الأساسية للبنية المحورية هي FlgG للقضيب و FlgE للخطاف و Flic لليف و كلها تتجمع بمساعدة بروتين cap (FlgJ و FlgD و FliD على التوالي). و منها كلها يبقى فقط FliD عند طرف الليف في الناتج النهائي. هناك مكونات أخرى في البنية المحورية (تسمى FlgB و FlgC و FlgF) تصل القضيب و مركب الحلقات MS. فالخطاف و الليف يتصلون عبر FlgK و FlgL. عندما تثبت الحلقة C و القضيب C في الحلقة M في سطحها السيتوبلازمي فمركب الحلقات MS–الذي يعتبر الأساس البنيوي للجهاز- يمكنه البدأ بإفراز البروتينات السوطية.
فبنية القضيب تتشكل عبر طبقة
البيبتيدو غليكان، لكن نموها لا يستطيع اجتياز الحاجز الفيزيائي المقدم من
طرف الغشاء الخارجي دون أدنى مساعدة. لذلك فإن مركب الحلقات يحفر حفرة في
الغشاء و هكذا يستطيع الخطاف النمو بفضل تنسيق FlgD حتى يبلغ الطول المحرج 55 nm. و الركائز التي تتشكل تتحول من وضع خطاف-قضيب إلى وضع فلاجيلين، و FlgD يمكن أن يستبدل بروتينات مرتبطة بالخطاف و الليف يستمر في تشكله. فمن دون البروتين cap Fli Dفمونومرات الفلاجلينسوف تضيع و بالتالي فإن البروتين capيعتبر ضروريا لكي تتم العملية.
تطور السوط انطلاقا من نظام الإفراز من النمط الثالث (TTSS) لا يعمل:
يمكن أن يفكر أحد ما أن الوصف المقدم أعلاه يمكن أن يكون أكثر من كاف للضرب بعرض الحائط التفاهات التي قدمها كين ميلر (Kenneth Miller)
و آخرون إلا أن الوضع يزداد سوءا أكثر للتاريخ الدارويني. لماذا التخليق
الحيوي للسوط بالضبط هو هكذا انضباطا و تناسقا؟ ليس فقط لأن متطلبات الطاقة
تجعل من السوط نظاما جد مكلف و لكن لأن التعبير المبكر للبروتينات السوطية
ربما يؤدي إلى استجابة مناعية قوية في النظام المستضيف و هو ما لا تود
البكتيريا فعله.
ما المغزى من هذا الأمر من وجهة
نظر تطورية؟ حسنا، فمونومرات الفلاجلين هي إلى حد ما محرضات سيتوكين قوية.
إذا ما كنت أنت عبارة عن ييرسينيا (Yersinia)
فبامتلاك نظام الإفراز من النمط الثالث فآخر شيء ستفعله هو إظهار بيبتيدات
الفلاجلين للبلعميات الكبيرة. فشيء كهذا فلا شك من أنه سيكون ضارا للآليات
المضادة للالتهابات الموجودة في الييرسينيا.
الخاتمة:
إن الوصف الذي قدمته أعلاه لم يمس سوى سطح هذا الموضوع الرائع من تكنولوجيا النانو (للمزيد من المعلومات انظر هنا).
و بهدف الإيجاز فأنا لم أناقش بعد بخصوص عملية الانجذاب الكيميائي و عنصري
نقل إشارة مجموعة الدوائر الكهربائية و التحويل الدوراني و قوة البروتون
التي تؤدي إلى اشتغال السوط (لتفاصيل أكثر أنظر هنا).
لكن بيت القصيد هنا هو أن نظرية الداروينية الحديثة- كما تفهم عادة- لم
تعطي أي تفسير بخصوص أصل هذه الآلة الدافعة و المعقدة بشكل ملحوظ. ربما كما
حدث مع التفسيرات الداروينية بخصوص العين و التي في البدء تبدوا أن
بإمكانها إقناع المبتدئين الغير المتعودين على روائع الكيمياء الحيوية و
على الأساسات الجزيئية للرؤية. و هكذا فسرعان ما تصير هذه التفسيرات
الداروينية فارغة و غير مقنعة عند الأخذ بالاعتبار التفاصيل الجزيئية
للنظام. و عندما يجمع شخص بين التفاصيل المقدمة في الأعلى و بين العجز
الكلي للداروينيون الجدد عن إنتاج طيات جديدة من البروتينات و مواقع جديدة
من الاتحاد البروتيني-البروتيني، حينها يظهر السؤال التالي: هل يمكن لهذا
النظام أن ينسج نفسه عن طريق تغيرات طفيفة و متوالية و عبر مراحل؟ فبما أن
الاقتراح الأساسي للداروينية يخدع بقدرته المزعومة على إعطاء تفسير لوفرة
البراهين الدالة على التصميم. ألا تبدي الآن عجزها الواضح عن دحض مسلمة
التصميم المشار إليه من قبل كمقترح علمي واقعي و محترم؟
دوغلاس أكس (Douglas Axe) من المعهد البيولوجي (Biologic Institute) أظهر في بحث حديث قد تم نشره في مجلة Bio-complexity
أن نموذج التضاعف الجيني و التوظيف يعمل فقط عندما تتطلب تغيرات صغيرة
لاكتساب فائدة أو وظيفة جديدتين قابلتين للانتقاء. إذا كان الجين المضاعف
محايدا (من حيث تكلفتها بالنسبة للكائن الحي) فالعدد الأقصى للطفرات الذي
يتطلبه أي تجديد داخل مجموعة من البكتيريا فهو أكثر من ستة. و إذا كان
الجين المضاعف ذو تكلفة سلبية بشكل طفيف فالعدد الأقصى سيتراجع حينها إلى
اثنين أو أقل (دون إدراج الضعف نفسه).
يبدو أن السوط البكتيري يعتبر
–ربما بشكل مذهل- تحديا حقيقيا للتطور الدرويني مثل ما كان عليه الحال منذ
1996 عندما ذكره بيهي لأول مرة في كتابه: "الصندوق الأسود لداروين".
التعريف بالكاتب:
جوناتهان ماكلاتشي (Jonathan McLatchie) حاصل على ماجستير البحوث في البيولوجيا التطورية و النظامية في جامعة كلاسكوو Glasgow. حاليا هو محرر موقع "التطور أخبار و نظرات
فديوهات توضح كيف يعمل السوط البكتيرى وهل يمكن اختزاله
هذا الفديو يظهر كيف سياط البكتيرية تبدل و تغير اتجاها و دورانها
للاستجابة للمؤثرات الكيميائية؟ و يظهر ان حتي طريقة تحركه سواء عكس عقارب
الساعة او مع عقارب الساعة له بروتين خاص به كي تستجيب للمؤثرات الكميائيه و
يتحكم في















اروع شي قرائته عندك محمد
ردحذفلك موتور موتور كهربائي هذا تصميم مش ممكن
ردحذفهل منهم من يعقل ذلك
حذفلا اعتقد :)