الجينات الزائفة
مقدمة و رؤس اقلام
.......( pseudogenes ) .......
الجينات الزائفه دليل زائف أخر على التطور
الجينات الزائفة هو مصطلح اطلق على تسلسلات من الماده الوراثيه التي تشبه
الجينات الوظيفية ووكان الاعتقاد السائد انها لا تقوم باى غرض وظيفى
معروف وهى مجرد بقايا وركام لأجزاء جينية غير قادرة على أداء وظيفتها
بسبب أضرار مميتة أصابت بنيتها و الغالبية العظمى منها مجرد نسخ مصابة
بأذى لجينات وظيفيه عاملة،
استخدم انصار التطور الجينات الزائفه كمدلول لركام او حطام تالف لجينات حيه
وظيفيه كدليل تطورى واعتبر من اقوى دلائل التطور والسلف المشترك
باعتبارها بقايا لجينات ماتت منذ زمن بعيد واستخدمت تلك الجينات الزائفه
كأحافير (مستحاثات) جينية،وراثيه ..... وتم تفسير اشتراك تلك الجينات
التالفه والمعطله بين انواع الاحياء المختلفه كدليل حتمى على اشتراك السلف
والنسب التطورى باعتبار ان تلك الاخطاء لا يمكن ان تحدث بطريق الصدفه
بنفس المناطق الجينيه مع كونها ركام وبقايا غير نافعه وليس لا اى فائده
للكائنات الحيه الحامله لها ولا معنى لوجودها بدون وظيفه ....
ومن
اشهر الامثله التى يكررها انصار التطور كدلاله حتميه على اشتراك البشر
والشامبنزى فى سلف قديم هى الجينه المعطله .Gulo. التى تصنع إنزيما هو
العنصر الأخير في السبيل الكيميائي الحيوي لتركيب الفيتامين C. واعتمدوا
كونها معطله فى الانسان والشامبنزى دلاله على سلف مشترك
.
لكن ....
ماذا لو كانت هذه الجينات غير زائفه وليست مجرد ركام وتؤدى وظائف حيويه وتنظيميه هامه ؟....
....................................................
بالطبع لن يكون لاستدلال التطوريون بها اى معنى حيث ان اثبات وظيفتها
يدحض تلك الحجه ويبرر وجودها بكيانات الكائنات الحيه ولم تعد تلك
الحفريات الوراثيه المتناثره داخل اجسادنا ...
تتوالى الدراسات
والابحاث التى تؤكد تلك الحقيقه منذ بداية القرن حول وظيفة الجينات
الخادعة ودورها في تنظيم فاعلية الجينات الوظيفية..... ففي السنوات
الأخيرة، توصل علماء البيولوجيا الجزيئية إلى إدراك أن كثيرا من الجينات في
الكائنات الحية العليا لا تكود نتاجا پروتينيا نهائيا، بل عوضا عن ذلك
تعمل نسخ الرنا الخاصة بهذه الجينات على ضبط فاعلية جينات أخرى. وبإمكان
هذه الجزيئات المنظمة من الرنا .(regulatory RNA molecules ).أن تُفَعِّل
أو تكبت على نحو متباين جينة أخرى، أو أنها تتدخل في ترجمة نسخة الرنا
المرسال لتلك الجينة إلى پروتين وظيفي ......ففى عام 1999 كانت نتائج
لدراسه من قبل مجموعة. <M.أوشيا> [البحثية من جامعة سوسكس
بإنكلترا]؛ إذ أوجد الباحثون في عصبونات حلزون البركة الشائع..(common pond
snail).. أن كلا من الجينة لسينثاز أكسيد الآزوت ..(nitricoxide synthase
(NOS ..وقريبتها الجينة الزائفه (كما كان يعتقد) تنتسخان إلى رنا، ولكن
نسخة الرنا للجينة الخادعة .NOS .تثبط إنتاج الپروتين من قبل نسخة الجينة
السوية NOS.....
دراسات موسعه عن الدور التنظيمى الحيوى فى عام 2002 وعام 2003
http://www.sciencedaily.com/releases/2002/08/020830072103.htm
http://www.ncbi.nlm.nih.gov/pubmed/14616058
دراسه اخرى في عام 2003.، بواسطة <S.هيروتسون> [من كلية طب سيتاما
في اليابان] حيث تتبع أثر التشوهات في مجموعة من صغار فئران التجارب في ما
يتعلق بتبدل جينة خادعة
.وتوصل لان سبب تلك التشوهات هو إبطال فاعلية جينة منظمة مهمة تعرف بماكورين Makorin. .
لكن <هيروتسون> لم يمس الجينة الاصليه بشيء، بل عطل مصادفة الجينة
الخادعة لتلك الجينه الاصليه والتى اطلق عليها Makorin1-p1.. ، وهذا أثَّر
في وظيفة نظيرتها الجينة ماكورين
http://www.ncbi.nlm.nih.gov/pubmed/12721631
http://www.nature.com/nature/journal/v423/n6935/full/nature01535.html
لكن الامر اللذى قلب موازين التعريف للجينات الكاذبه هى دراسات اثبتت ان
تلك الجينات جينات كامنه تنشط وتقوم بوظائفها حين تواتيها ظروف بيئيه
مختلفه كما اوضحت احدى الدراسات على الخميره حيث يتم إعادة تفعيل جينات
خادعة لپروتينات خلوية السطح معينة وذلك عندما يُجابه الكائن الحي ببيئة
جديدة
دراسات حديثه ايضا تحدد بدقه وظائف بعض الجينات الكاذبه
وتاثيرها بالتوازن الذي يؤثر على مستويات التعبير الجينى كالجين PTENP1
وتاثيره المباشر على ظهور بعض الامراض ك..Bannayan-Zonanal ..وغيرها من
الامراض
http://www.wired.com/wiredscience/2010/06/expanding-genome
دراسات واوارق بحثيه متعدده للامثله لا للحصر توضح الدور الحيوى البالغ الاهميه للجينات الزائفه
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2981145/
http://www.nature.com/nature/journal/v453/n7194/full/nature06904.html
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3206313/
الابحاث تتوالى تباعا لتحطم امالا لانصار التطور وتهدم قلعه رمليه اخرى
حول الجينات الزائفه ..( pseudogenes ) التى اسموها الحفريات الجينيه
الداله على الاسلاف المشتركه
وكان الدليل الوحيد هو الجهل بمعرفة وظيفتها
ونظرة عن كثب
اول الجينات المزيفة تم الاخبار عنه عام 1977م منذ ذلك الوقت، تم
الإبلاغ عن عدد كبير من هذه الجينات في البشر، ووصف والعديد من الأنواع
الأخرى.
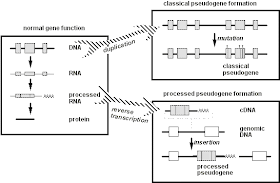
هناك نوعان من الجينات الزائفة"المعالجة " و"غير المعالجة ""
الجينات الزائفة المعالجة موجودة على كروموسومات مختلفة عن نظرائها
الفاعلة , أنها تفتقر الإنترونات وجينات منظمة معينة، وغالبا ما تنتهي في
سلسلة الأدينين، ومحاطة بتكرار المباشر (التي ترتبط مع العناصر الجينية
المتحركة). قد تكون نسخ كاملة او غير كاملة من احد الجينات أو خليط من عدة
جينات. ويعتقد أنها نتجت خلال عملية من 3 خطوات الاولى هي نسخ ال DNA الى
RNA
الثانية تغيير الانترون الى mRNA
والثالثة تحويل mRNA الى DNA مرة اخرى عن طريق النسخ العكسي .
هذه الطريقة يعتقد انها انتجت عائلة L1 من الجينات الزائفة
نظرية اخرى تقول ان الجينات الزائفة جائت من فيروس النسخ العكسي الداخلي التي انتقلت بين الانواع المختلفة .
الجينات المزيفة غير المعالجة عادة ما تكون موجودة في تكتلات بنفس
التسلسل الوظيفي على نفس الكروموسوم.غالبا ما تكون تمتلك انرونات وما
يرتبط بها من جين منظم . التعبير عنها عادة ما يكون ممنوع من قبل كودون او
كودونات ايقاف موضوعة في مكان خاطئ.قد تكون هنالك تغييرات اخرى كنتيجة
لالحذف او الادخال وطفرات نقطية .بعض انواع ال mRNA قد او قد لا يتم
انتاجها اعتمادا على نتيجة الدمار في الجين .
ويعتقد أن العديد من هذه قد نشأ من مضاعفة الجينات التي تنتج نسخة
إضافية من الجين. يمكن للنسخة الإضافية ان تراكم الطفرات دون التاثير على
الكائن الحي لان النسخة الاصلية لم تتعرض للطفرات
( فرضية التضاعف الجيني التطورية تقترح انه يمكن بمرور الوقت ومن خلال
طفرات عشوائية ايجاد جين جديد بوظيفة جديدة باستعمال هذه النسخة المضاعفة
مع الاحتفاظ باوظيفة النسخة الاصلية طفرات عشوائية ايجاد جين جديد بوظيفة
جديدة باستعمال هذه النسخة المضاعفة مع الاحتفاظ باوظيفة النسخة الاصلية )
الجينات الزائفة المشتركة
ويرى تطوريون أن الجينات الزائفة المشتركة،
التي ليس لها وظيفة بأي شكل من الأشكال في أنواع مختلفة، انها من الأمثلة
على أصل مشترك. مقارنة تسلسل الحمض النووي من البشر، الشمبانزي، وغيرها من
الثدييات يظهر عدد كبير من الجينات الزائفة المشتركة. ولعل المثال الأكثر
شهرة لجين زائف مشترك هو جين ايتا-غلوبين.
ويقع هذا الجين على الكروموسوم ايتا 11 في البشر والرابع في سلسلة من 6
جينات غلوبين بيتا (خمسة منها فقط عاملة) .4 ليس لديها أي كودون بداية (
AUG ولها العديد من كودونات الايقاف. من الواضح جدا، انه ليس هناك أيةmRNA
وبالتالي لا بروتينات تصنع. البشر، الشمبانزي، والغوريلا لها نفس العدد
من الجينات بيتا غلوبين مرتبة في تسلسل نفسه. تسلسل الاكزونات في هذه
الجينات هي أيضا مشابهة - كما هو الحال مع اكزونات الجين ايتا
ويغتقد ان الجين ايتا غلوبين نشا من مضاعفة الجين غاما غلوبين بسبب
التشابه العالي بين تسلسلات الجينين كما ان كلا الجينين موجودين في
الرئيسيات .
ويعتقد أن الجين الزائف ايتا غلوبين قد نشأ نحو 140 مليون سنة مضت في
الجرابيات والثدييات المشيمية. بعد "الانفصال التطوري" للجرابيات تم
تشكيل، الجين غاما غلوبين من مضاعفة احد الجينات في اسرة البيتا غلوبين.
في وقت لاحق، ولكن قبل تشعب رتب الثدييات المشيمة، الجين ايتا-غلوبين شكل
من مضاعفة الجين غاما غلوبين.بالتالي يتوجب وجود الجينات
جاما و ايتا في
الأجداد ذوات المشيمة ولكن وبصورة افتراضية قد فقدت غاما من الماعز
(التي ليس لديها غاما) وفقدت ايتا من الأرانب (اللتي لم يكن لديك ايتا).
وفقا لهذا السيناريو، يجب أن يكون قد الجين ايتا كان عاملا في البداية،
لأنه هو وظيفي في الماعز اليوم. 2 وغير عامل في جميع الرئيسات، الامر
الذي يتم تفسيره على أنه يعني أنه كان بالفعل غير فاعل في القرود الأجداد
70-80 مليون سنة. هذا التفسير يعني أنه قد تم الحفاظ على الجينات
غلوبين-ايتا لأكثر من 70 مليون سنة دون أن تحويلها إلى جينات جديدة مفيدة
ودون القضاء عليها من خلال الطفرات العشوائية
علامات الوظيفة؟
لذا، فإن استمرار وجود تسلسل الحمض النووي غير فاعل في النسب كامل
للجينات كل هذه الفترة الطويلة من الوقت المفترض يبدو ملحوظا في سياق
فرضية مضاعفة الجينات. وحقيقة أن الجينات الزائفة لا تزال موجودة ووتتم
تمييزها بعد عشرات الملايين من السنين دون أي وظيفة مفيدة لا يبدو أن له
معنى. بالتأكيد، فانه من دون وظيفة نافعة، فان الانتقاء الطبيعي ما كان
ليحافظ على تسلسل لفترات طويلة من الوقت مثل هذه. هناك في الواقع تكلفة
للحفاظ على
DNA. غير فاعل فإنه يأخذ الطاقة لتكرارنفسه والحفاظ على الحمض النووي
الذي لا يدفع بالمقابل للحفاظ على أعماله. على الرغم من أن هذا قد يبدو
ضئيلا على المدى القصير. حتى بتكلفة ضئيلة للغاية تتفاقم على مدى الملايين
من الأجيال يبدأ بالتحول إلى ضرر كبير. لذلك، فإن حقيقة أن لديك جينات
زائفة مميزة وتمتلك تركيب شبيه بالجينات يؤدي الى الاستنتاج بانها فعلا
تؤدي نوعا ما من الخدمة .
استمرار بقاء الجينات الزائفة هو في حد ذاته دليل على نشاطها. هذه
مشكلة خطيرة بالنسبة للتطور، حيث من المتوقع أن الانتقاء الطبيعي من شأنه
أن يزيل هذا النوع من الحمض النووي لو كانت عديمة الفائدة، منذ
DNA المصنعة من قبل الخلية مكلفةمن ناحية استهلاك الطاقة. بسبب عدم وجود ضغط انتقائي على هذا الحمض النووي المحايد(neutral DNA)
، فإن للمرء أن يتوقع أن الجينات الزائفة"القديمة" كان يصعب التعرف
عليها نتيجة لتراكم الطفرات العشوائية. وعلاوة على فان الية ازالة الجمض
النووي المحايد الان باتت معروفة

"عندما يقول الناس عادة أن الجينوم البشري يحتوي على 27000 جين أو نحو
ذلك، فإنهم يشيرون إلى الجينات التي تشفر للبروتينات،"كما يشير جورج
ميشيل، استاذ في علم الوراثة في جامعة لييج في بلجيكا. ولكن على الرغم من
هذا العدد هو ما يزال في بدايته،( تتراوح التقديرات من 20،000 إلى 40،000
)يبدو ان هذا العدد يؤكد عدم وجود علاقة واضحة بين تعقيد النوع وعدد
الجينات في جينومه. "ذباب ثمار الفاكهة لديها أقل من جينت التشفير في
الدودة المستديرة، ونبات الأرز لديها أكثر من البشر"، ويلاحظ جوناس ماتيك،
مدير معهد العلوم البيولوجية الجزيئية لفي جامعة كوينزلاند في بريسبان،
أستراليا. "ان كمية الحمض النووي غير المشفر تتناسب مع تعقيد الكائن الحي
". ...
"نحن على نحو متزايد ندرك أن هناك مجموعة كبيرة من" الجينات "التي هي
وظيفية بصورة واضحة على الرغم من أنها لا تشفر أي بروتين" لكن تنتج فقط ار
ان اي. يشير جورج الى ان مصطلح "جين" كانت دائما فضفاضا وان هذه ال ارن اي
(رنا )هي فقط جينات تبحث عن معناها بصورة ابعد .لتجنب الارباك يقول كلاس
والشتيد
من معهد كارولينسكا في السويد، "نحن لا نميل إلى الحديث عن 'الجينات'
بعد الآن؛ نحن نشير فقط إلى أن أي جزء هو ما كتب [إلى رنا] على أنه 'وحدة
النسخي.' بناء على مسح تفصيلي لجينوم الفأر لجميع هذه العناصر، وقال" نقدر
أنه سيكون هناك 70،000 إلى 100،000،من هذه العناصر "أعلن والشتد,
في المؤتمر الدولي لعلم الوراثة، الذي عقد في يوليو الماضي في ملبورن."
نصف هذه العناصر بسهولة يمكن أن تكون غير مشفرة. "إذا كان هذا هو الصحيح،
اذا لكل تسلسل الحمض النووي الذي يولد البروتين، فان هنالك آخر يعمل فقط
من خلال الأشكال النشطة من رنا
-أشكال مخططات التي ليست مجرد وسيط للبروتينات، بل يغير مباشرة سلوك الخلايا ". ...
"يقول ماتيك "أعتقد أن هذا سوف يكون قصة كلاسيكية كيف تقوم العقيدة
بعرقلة التحليل الموضوعي للحقائق، في هذه الحالة لمدة ربع قرن،" .
ان الفشل في تحديد هذه الامكانية المفترضة في ان تكون هذه المقاطع غير
المشفرة ممكن ان تنقل معلومات بصورة جزيئات رنا سيعتبر اكبر خطا في تاريخ
الاحياء الجزيئية
ونظرا لهذا، فمن غير المعروف ما إذا كان كل الجينات الزائفة المعروفة
حاليا هي فعلا بلا وظيفة. في الواقع، يعتقد ان بعض الجينات الزائفة تعمل
كمصدر للمعلومات لإنتاج التنوع الجيني. ويعتقد أن الجينات الزائفة الجزئية
يتم نسخهاالى نوع من الجينات الفاعلة خلال إعادة التركيب الجيني. وقد تم
الإبلاغ عن هذه الظاهرة مرات عديدة لتشمل البروتينات المناعية المختلفة
داخل الفئران والطيور والجينات هيستون الفار، جينات غلوبين الحصان،
والجينات البشرية غلوبين بيتا. ومن غير المعروف ما إذا كان هذا يمكن أن
يكون الدور الذي يمكن لهذا الجين غلوبين-ايتا كذلك. ومع ذلك، فإن حقيقة أن
يقع الجين الزائف ايتا-غلوبين بين الجينات الجنينية وجينات البالغين يشير
إلى أنه قد يلعب دورا في التحول الجيني (يبدو أن هناك بعض الأدلة الأولية
لهذا الغرض على الرغم من أن جزءا من تسلسل الجينات ايتا لا يزال مجهولا
).
يبدو ان الجزء الذي يعطي الشفرات للروتين هو معلوماتيا مبسط ويبدو ان
التعقيد المعلوماتي يقع في الجزء غير المشفر هذا الجزء من الجينوم يوجه
متى وأين يتم وضع لبنات بناء البروتين، وبالتالي في غاية الاهمية لتحديد
الهيكل العام والوظيفة النهائية للمخلوق الناتجة عن ذلك. و بسبب التحيز
التطوري ظلت هذه المناطق من الحمض النووي تعتبر خردة لفترة طويلة من الزمن
- وبالتالي تم التغاضي عنها ولم يتم تمييزها بوصفها عناصر معلوماتية
رئيسية في الجينوم. ومن المثير للاهتمام، هذه النتائج تدعم فعلا تنبؤات
نظرية التصميم الذكي في سياق مكافحة منذ فترة طويلة الافتراضات التطورية.
بالطبع، هناك دائما تعديلات مغرضة لشرح مثل هذه التنبؤات الفاشلة الناجمة
عن التحيز التطورية.

الإنسان مثل شقائق النعمان البحر؟
دعم هذا فكرة الطوب والبلاط هو مضمون دراسة أجريت عام 2007 نشرت في
مجلة العلوم من قبل بوتنام واخرون. على الجينوم المثير للاهتمام لشقائق
البحر .في هذه الورقة البحثية دون الكاتب ان المظهر العام لجينوم شقائق
النعمان البحرية هذه هو شبيه الى حد كبير بجينوم الانسان ...نعم جينوم
الانسان.
"واحدة من المفاجآت الكبيرة لجينوم شقائق النعمان، يقول سوالا، هو
اكتشاف كتل من الحمض النووي التي لديها نفس جينات التكملة كما في الجينوم
البشري. الجينات الفردية قد تبادلت الأماكن، ولكنها غالبا ما ظلت مرتبطة
معا على الرغم من مئات الملايين من السنين من التطور ... وعلاوة على ذلك،
تبدو جينات شقائق النعمان شبيهة بالفقريات وهي غالبا ما تكون مليئة بمناطق
لا تعطي اي شفرة تدعى انترون.
، والتي هي أقل شيوعا بكثير في الديدان الخيطية وذبابة الفاكهةعنها في
الفقاريات من. وأكثر من 80٪ من الإنترونات في شقائق النعمان في نفس
الأماكن في جينوم البشر ...
فينيرتي وطالب الدراسات العليا له سوليفان جيمس بدا أيضا فيالنظر الى
جينوم شقائق النعمان ل283من الجينات البشرية المشاركة في مجموعة واسعة من
الأمراض. وقالوا في عدد يوليو من الخريطة الوراثية البشرية انهم وجدوا
226. وعلاوة
على ذلك فان جين سرطان الثدي هو نفسه بين الانسان وشقائق النعمان عنه في الانسان والديدان الشريطية او ذبابة الفاكهة
هذا يفرض بان حتى الجينومات القديمة كانت معقدة وتحتوي على الجينات اللازمة لبناء الكائنات متعددة الخلايا المعقدة الموجودة اليوم
لا يمكننا استبعاد احتمال، ،أن هذه الإنترونات المتخصصة ظاهريا كانت
موجودة في الجد المشترك الأخير من لكل من النباتات والفطريات والحيوانات،
ولكن ضاعت
بصورة متوازية في كل من النباتات والفطريات. . .
من اين اتت الكائنات البعديات الحقيقية (
eumeatzoan)
بذخيرتها ؟؟؟ حوالي 80 % (اي 6128 من اصل 7766)من جينات اسلام البعديات
الحقيقية لها اقارب معروفين (ما معناه بروتينات مع تطابق نسبي في التسلسل
وتركيب هيكلي محفوظ ) من خارج مملكة الحيوان من النباتات والفطريات
وانواع اخرى متوفرة من النشر العام للبيانات . هذه بصورة اكيدة جزء من
جينات حقيقة النواة القديمة التي كانت موجودة في السلف وحيد الخلية الاخير
للكائنات متعددة الخلايا وهي تدخل في صلب عمل الخلايا حقيقة النواة
كما ذكر أعلاه، فإن هذه الورقة تدعم فكرة أن الجينات داخل الجينوم
الأساسية هي اللبنات بسيطة جدا والتي يمكن أن تكون هي في الواقع تستخدم
لبناء العديد من أنواع مختلفة من المخلوقات - من البشر إلى شقائق النعمان
للنباتات والفطريات، وحتى واحد خلايا الكائنات الحية مثل الهدبيات .
وباختصار فان فانه ليست المناطق المشفرة للبروتين هي المسؤولة عن الصفات
الظاهرية للكائن الحي بل ان المعلومات اللازمة لتركيب وتجميع الكائن الحي
موجودة في المناطق غير المشفرة اذ . اذ انه يمكن استدام اللبنات الاساسية
كالطوب لبناء غرفة واحة او بناء ناطحة سحاب كل ما نحتاج له هو مخططات
مختلفة لتوجيه تلك اللبنات .
ورقة بوتنام ايضا تعارض الفكرة القائلة بانه ممكن ان نرسم اشجار وراثية
بناء على اسس جينية شكلية بصورة مستمرة . ولكن بعد كل هذا يظهر ان شقائق
النعمان تمتلك تجانس في الجينوم مع الانسان لا تظهره مع ذبابة الفاكهة او
الديديان الشريطية فكيف يمكن تبرير هذا من دون استضافة دعامات مخصصة من
قبل العلماء الدارونيين ؟؟؟ في الواقع ان الاحتجاج بان هذه الجينات قد
ضاعت من قبل بعض الانواع واحتفظ بها في انواع اخرى على مدى بلايين السنين
يبدو غير منطقيا البتة (او على الاقل هكذا يبدو لي ).
خردة واحدة للانسان
مجموعة اخرى من الجينات الزائفة الموجودة في الانسان وتدعى ترانبوسونات
مثل ( عناصر الو ) والتي كان يعتقد انها غير ذات فائدة تضح انها لديها
وظيفة مهمة .
هنالك ادلة متزايدة على ان تسلسلات الو (عناصر نووية قصيرة ومتخللة )
تلعب دورا في تنظيم عمل الجين ممكن ان تنشطه او تثبطه كما انها تعمل
كمستلمات (مواضع التحام ) وهذا ممكن يكون مقدمة لبيان عمل الجينات الزائفة
الاخرى
في عام 1997 نشر فلام ومجموعته بحثا في مجلة العلوم يقول فيه ان الحمض
النووي الخردة مرتب مثل ترتيب اللغة البشرية ونشر ادلة لغوية تدعم ادعائه.
العلماء فسروا هذا التشابه على انه رسالة مضمنة في الحمض النووي الخردة
رغم انها ستظل لغز يمكن ان يحله كل انسان بطريقته .هذه المحاججة مهمة
لانها ممكن ان تستخدم مثلا لاثبات وجود ذكاء لكائنات خارجية (مثل اي تي )
لو تم العثور على تركيب شبيه باللغة مثلها عن طريق التقاط موجات صوتية او
وجدت محفورة على الصخور المتساقطة من السماء
حوالي عام 1998 بدأ كارل شميت، عالم الأحياء الجزيئية في جامعة
كاليفورنيا في ديفيز، النهوض بما بدا وكأنه فكرة مجنونة لشرح الالفة غير
الاعتيادية بالنسبة لألو للجينات. اقترح شميد بان وجود التسلسلات الو في
مواقع قريبة عن الجينات ممكن انه يعني ان لها دورا في اصلاح الجينات
وصيانتها . وبوجود خريطة جينوم الانسان امامهم والتي تظهر وجود الكثير من
تسلسلات الو حول الجينات بدا العلماء اخذ كلام شميد على محمل الجد حيث قال
العالم فرانسيس كولينز "ان هذا يبدو لطيفا " وقال اريك لاندر انه يوافق
وفي الآونة الأخيرة في عام 2001، اكتشف فريق من علماء الوراثة الجزيئية
اثنين من"النقاط الساخنة" حيث نفس العناصر النووية القصيرة المتخللة
(سينس) تم إداخلها في الجينوم بشكل مستقل:
وقد استخدمت الترانسبونات العكسية للفقاريات على نطاق واسع لتحاليل
ودراسات الجينية والشكليةوالتطور الجزيئي. يمكن الحصول على معلومات محددة
من دراسة ادخالات معينة في تسلسل الحمض النووي إما من خلال مقارنة
الاختلافات التسلسلات هذه الادخالات التي تراكمت على مر الزمن من خلال
مقارنتها مع نسخ مناظرة لهذه العناصر المحددة او من خلال تحديد وجود او
غياب هذه العناصر في موقع معين. فقد اعتبر وجود نسخ محددة لتكون بشكل
اساسي ذات طابع النشوء والتطور حيث ان نسبة احتمال وجود اكثر من ادخال يتم
بصورة مستقلة في مكان واحد هي صفر.لقد تم تحديد نقطتين ساخنتين لادخال
العناصر النووية المنتشرة القصيرة SINE في داخل منطقة mys-9 وفي كل منطقة
ساخنة وجد اثنان من ادخالات العناصر النووية المنتشرة القصيرة في مواقع
متطابقة . هذه النتائج كان لها انعكاس كبير على تحليل نشوء التطور
بالاستناد الى العناصر النووية ( SINE )
مما يشير الى ضرورة توخي الحذر عندما يتم الاستنتاج بان وجود العناصر النووية ساين يدل على وجود السلف المشترك.
حتى الآونة الأخيرة، نشرت جيني لي في عدد مايو 2003 من الطبيعة، مقالا
بعنوان "التواطؤ من الجينات والجينات المزيفة" التي قدمت بعض النتائج
المثيرة للاهتمام من العمل الذي قام به هيروتسون واخرون.:
بكونها غير مفيدة في انتاج البروتينات فان الجينات الزائفة قد تم
استبعادها كجينات فاعلة ولكن السؤال لماذا حافظت الثدييات على تراكمها
لهذا الحد الكبير؟ احد الاجوبة المقترحة هو انه بارغم من كون الجينات
الزائفة تؤدي كاثار تطورية وازعاج للتحليل الجيني فان الطريقة التي تحتاج
للنشوء من خلالها تنشئ اسرا جينية كاملة مثل تلك التي تشترك في عملية
المناعة او حاسة الشم لكن، هل الجينات الزائفة أنفسهم مجرد تركات لهذه
العملية؟ أو هل ان القيام ب(كالانتقاء الطبيعي ) للاحتفاظ بها. إشارة الى
وجود وظيفة بيولوجية خفية لديها ؟ ، وهذا الادعاء الأخير يبدو أن يكون
صحيحا.هيروتسون وزملائهة قدموا نتيجة غير مسبوقة في هذا المجال تفيد ان
الجين الزائف Makorin1-p1 الذي يقع على الكروموسوم 5 في الفئران يلعب
دورا في حيويا مهما (يقوم بتنظيم انتاج الجين
Makorin1-p1(الذي يقع على كروموسوم مختلف تماما – الكروموسوم 6 في الفئران)
ان نتائج عمل هيروتسون وزملائه هي محفزة للكشف عن وظيفة الجينات
الزائفة التي كانت تعد ليست اكثر من مجرد متحجرات جزيئية (صور لتجارب
الطبيعة الام السابقة التي مضت )
في مقالةجديدة لمجلة العلوم قام ووجشيش ماكالاوسكي بتعليق يبدو انه يتناغم مع ما كان علماء نظرية التصميم يرددونه منذ زمن :
على الرغم من جاذبيته فان مصطلح (الحمض النووي الخردة ) قد ادى الى
نفرة تيار كبير من العلماء عن دراسة الحمض النووي الذي لا يشفر البروتينات
. ولكن من كان ليتوقع ان مجموعة صغيرة من العلماء الجينومينن قد بحثت في
القمامة هذه ؟ ففي العلم كما في الطبيعة فان هنالك مجموعة من الناس مستعدة
للمخاطرة بان تبدو سخيفة في سبيل استكشاف اراضي جديدة وبسبب عملهم فان
النظرة عن الجينات الزائفة وخصوصا العناصر المتكررة قد تغيرت من عام اوئل
التسعينات . والان المزيد والمزيد من العلماء بدات تنظر للعناصر المتكررة
ككنز جينومي.
وفي ديسيمبر من عام 2003 م وفي العدد السنوي من "المراجعة السنوية لعلم
الجينات " نشر باراكيف وايالا ورقة بحثية بعنوان " الجينات الزائفة هل هي
احماض نووية خردة ام فاعلة ؟ " .وبالنظر لبعض من استنتاجاتهم سنكتشف انهم
يرددون ما يقوله علماء نظرية التصميم منذ فترة من ان الجينات الزائفة هي
فاعلة وهي في الواقع ليست زائفة بعد الان :
الجينات الزائفة تم تعريفها كونها مشتقة من جينان فاعلة وعلى هذا
الاساس فان الطفرات فيها ينبغي ان تكون محايدة ولها فرصة متساوية ان تكون
ثابتة في مجموع السكان.لكن الجينات الزائفة التي تم التحقيق فيها بشكل
مناسب في كثير من الأحيان تحمل أدوار وظيفية، مثل التعبير الجيني، تنظيم
الجينات، وتوليد التنوع الوراثي (الضد، المستضدية، وغيرها) . وتشارك في
تحويل الجينات الزائفة أو إعادة التركيب مع الجينات الوظيفية. كذلك تعاني
من الحفظ التطوري لتسلسلها وخفض تقلب النوكليوتيدات،وتغليب التنوع
النيوكليوتيدي النترادف على غير المترادف .وغيرها من المظاهر التي يتوقع
وجودها في الجينات او تسلسل الحمض النووي الذي لديه دور فاعل .. . .
شانها ان تضع الجينات الزائفة ضمن كونها ان وجود دراسات واسعة النطاق
وسريعة التزايد، لا يبرر وجود انقسام حاد خردة" تفتقر إلى وظيفة ولا تخضع
للانتقاء الطبيعي.وغالبا ما تكون الجينات الزائفة محافظ عليها ونشطة من
ناحية التعبير ..
يبدو ان هذه هي القضية الا وهي اكتشاف وجود فعاليةفي الجينات الزائفة
لكل الحالات او تقريبا للكل في حال وجود تحقق مناسب .المتابع سيستنتج
بسهولة هو انه الجينات الزائفة تمتلك او تحصل على نوع من الفعالية مما
يجعل من الخطا تعريفها كتسلسلات حمض نووي غير فاعلة تم الاحتفاظ بها من
جينات فاعلة او كجينات لا يتم التعبير عنها في الوقت الحالي لكنها تحمل
تشابه في التسلسل عن الجينات الفاعلة .بدلا من ذلك ينبغي تعريف الجينات
الزائفة على انها نتجت من مضاعفة او عكس المواقع للجينات الفاعلة والتي
غالبا ما تتعرض للانتقاء الطبيعي وبالتالي تحافظ على تسلسلها و تركيبها
الاصلي نظرا لانها اكتسبت ادوارا تنظيمية او ادوارا اخرى او ممكن ان تؤدي
خدمة كاحتياطي للتنوع الجيني
مطابقة الحمض النووي غير المرغوب فيه بين الإنسان والفأر -؟ الكثير من ذلك؟
ثم في مايو من عام 2004 م قام هاوسلر وبوجيرانو بمقرنة جينومات كل من
الانسان والفار والجرذ باستخدام الكمبيوتر , لقد كانوا يتوقعون بما انه
الانسان والفار الجرذ مختلفين عن بعضهم كثيرا فانهم سيجدون اختلافا جينيا
كبيرا .لقد وجدوا هذه الاختلافات المتوقعة في الجينات التي يتوقع انها
موروثة من السلف المشترك الا ان المفاجئة انهم وجدواامتدادات طويلة من من
الحمض النووي الخردة غير المشفر والتي تتطابق تماما بين الانسان والقوارض
"كان هناك حوالي خمسمائة امتداد من الحمض النووي في الجينوم البشري
التي لم تتغير على الاطلاق في الملايين والملايين من السنين التي تفصل
الإنسان عن الفئران والجرذان"، كما يقول هاوسلر. "أنني على وشك الوقوع من
كرسيي . انها مسالة غير عادية جدا أن يكون مثل هذا المبلغ من الحفظ
باستمرار على امتداد هذه الفترة الطويلة من الحمض النووي." 32
العديد من هذه الامتدادات من الاحماض النووية تدعى مناطق "عالية الحفظ
"، لا يبدو أنها تشفر للبروتين، لذلك قد يتم استبعادها كخردة إذا لم تكن
قد ظهرت في أنواع مختلفة كثيرة. هاوسلر اكد أن اختيار السلبي هو أقوى
ثلاث مرات في هذه المناطق مما هو عليه في المناطق غير المترادفة من
الاجزاء المشفرة . بقدر ما تشعر هاوسلر: "إنه لغزا كبير في تحديد الآليات
الجزيئية التي تضع تقريبا بصورة ماثلة للعيان كل قاعدة في قطعة من حجم
ما يصل الى 1 كيلو قاعدة [أي حوالي
يصل إلى 1000 زوج قاعدة ) تحت هذا المستوى من الانتقاء السلبي .تلك
المناطق من اكتدادات الحمض النووي والتي عددها 500 وتوطول الواحدة يصل الى
1000 زوج قاعدة هي متطابقة تماما في الانسان والجرذان – حوالي 500000
موقع حمض ننوي متطابق ما هو مثير للمفاجئة اكثر هو ان هذه المناطق تظهر
تطابق مع مناطق مماثلة في الدجاج والكلاب والسمك في حين انها غير موجودة
في ذبابة الفاكهة و بخاخ البحر . السلف المشترك الاخير لهذه الكائنات
يفترض انه عاش قبل 400 مليون سنة وبالطبع فان من المنطقي الاعتقاد بما انه
الطبيعة قد بذلت جهدا كبيرا للحفاظ على هذه الجينات عبر ملايين السنين
فانها لا يمكن ان تكون مجرد خردة كما يقول هاوسلر . بل هي تلعب دورا مها
في تنظيم نشاط الجينات التي لا غني عنها ونمو الاجنة ..
من خلال ما نعرفه عن معدل التغيير الجيني الذي يحصل في الحمض النووي
للبشر عبر الاجيال فان فرصة ايجاد امتداد حمض نووي لم يتغير بين البشر
والفئران عبر هذه المئات من ملايين السنين هي حوالي 1 تقسيم عشرة بجانبها
22 صفر ان هذا الكسر ضئيل ضئيل جدا بل واقعيا هو مستحيل ان يتم من خلال
الصدفة .
هذه المعلومات وضعت تحت الاضواء في التجرية التي اجراها نوبيرغا
ومجموعته والتي نشرت في مجلة الطبيعة عام 2004 م حيث اظهر وجود طفرات حذف
كبيرة في جينوم الفئران ( مجموعتين كبيرتين من الطفرات احداهما بطول 1511
كيلو قاعدة والاخرى بطول 845 كيلو فاعدة اي حوالي ما مجموعه 1,243000 زوج
قاعدة ) في الحمض النووي غير المشفر ممكن ان تحتملها الفئران دون تغيير
ملموس في فعاليتها .فئران قابلة للنمو ومتطابقة اللواقح تم ايجادها بعد
طفرات الحذف وكانت لا تفرق عن الفئران البرية في الشكل ,القابلية
التكاثرية , النمو , طول العمر , ومعايير اخرى تتعلق بالتوازن .ما هو مثير
للاهتمام هنا هو انه هذه المناطق غير المشفرة المحذوفة محفوظ بشكل عالي
في الانسان والفئران . يناقش الكاتب بان هذه المناطق غير المشفرة ممكن ان
تكون لها وظيفة لا تظهر على شاشتنا .مع ذلك فان ههذ الدراسة تدعم الفكرة
بوجود حمض نووي يمكن التخلص منه في جينوم الثدييات
السؤال هنا لماذا هذه الاحماض النووية اللتي يمكن التخلص منها محتفظ
بها لعششرات الملايين من السنين ؟ سواء ا كانت فاعلة ام غير فاعلة ؟ فلو
كانت فاعلة فهي اذا ليست بقايا التجربة والخطا لمحاولات الطبيعة الاولى .
ولو كانت غير فاعلة فما هو تفسير هذه الدرجة العالية من الحفاظ عليها ممكن
ان يتماشى مع فكرة ان السلف المشترك للانسان والقوارض (وكذلك الدجاج
والكلاب والسمك ) قد عاش قبل عشرات الملايين من السنين ).
وحتى مع ذلك، تلك مثل هاوسلر ونوبريغا ما زالوا يعتقدون بقوة أن البشر
والفئران قد انحدرا من سلف مشترك كان يعيش مئة سنة مليون سنة أو نحو ذلك.
فكرة أن البشر والفئران ربما قد تم إنشاء في الواقع بشكل فردي، عمدا، حتى
لا يعبر عقولهم.
وعلى اية حال فان هذه الاكتشافات يبدو على الاقل انها تدعم فكرة ان
جينومي الانسان والجرذان لا يقومان فقط بتشفير البروتينات ان حل لغز كون
هذه المناطق عالية الحفظ من وجهة نظر هاوسلر سيفتح الباب لاكتشاف اسباب
امراض مثل الصرع او التوحد حيث ان هنالك كثير من الامراض التي لا يصاحبها
تغييرات جينية وهذه مناطق جديدة ليتم البحث فيها .الاطباء الذين لديهم
عينات من الحمض النووي لمرضاهم ممكن يبحثوا في هذه العينات عن اي شئ مشترك
ممكن ان يشرح ما هو الشئ الخاطئ في مرضاهم – كيف يختلف الحمض النووي في
مرضاهم عنه في الناس الاعتياديين ؟ هذه المناطق هي اماكن عظيمة للنظر في
اسباب فيها عن اسباب امراض لا تزال مبهمة 32
هاوسلر يختتم بالجملة التالية "اعتقد ان هنالك نسبة من الحمض النووي
الخردة ستتحول لتكون ليست بخردة "اعتقد ان هذه هي قمة جبل الجليد وستكون
هنالك العديد من الاكتشافات المشابهة . بحلول عام 2007 هاوسلر وبوجيرانو
اكتشفا 10,402 تسلسل او ترانزبوسون والتي اظهرت فعالية يقول بوجيرانو "
اعتدنا على النظر اليها كمبعثرة للاشياء , ولكن هذه حالة تظهر فيها كذات
فعالية " . واعداد العناصر الفاعلة التي كان يعتقد انها خردة في تصاعد كبير
وبالاخذ بعين الاعتبار ماذا سيفعله التقاط هذه التسلسلات من تاثير على
الشجرة الوراثية . برايي لنت كون هناك اي شجرة وراثية بعد الان . ما اعنيه
ان وجود هذه التسلسلات بصورة متطابقة في الانسان والجرذان والدجاج
والكلاب سوف لن يجعل هنالك اي شئ مميز في وجودها متطابقة بين الانسان
والقردة .يبدو ان الصفات المشتركة سستحول لتكون صفات وظيفية محافظ عليها
جينيا اكثر من كونها روابط تطورية من سلف مشترك .
أخطاء مشتركة
حجة اخرى مثيرة للاهتمام هي انه الجينات الزائفة المتنوعة في انواع
مختلفة تمتلك اخطاء مشتركة .والتى لابد وانها نشات في سلف مشترك مع ذلك
فان هنالك ادلة على ان تسلسل الجينات هو ليس عشوائيا في بعض المواضع تم
التعرف على "نقاط ساخنة " طفرية في الجينات وكذلك في الجينات الزائفة .
طفرات نقطية بل وحتى انواع خاصة منها تم تحديدها وهي منتشرة بشكل كبير
اكثر من اي مكان اخر في الجين .
لناخذ على سبيل المثال الجين الزائف GULOPاو GULO في معظم الثدييات
فان هذا جين فاعل يرمز للللانزيم L-glucono-γ-lactone oxidase (LGGLO).
هذا الجين الزائف يقع في الكروموسوم 8 على الذراع p21.1
في منطقة غنية بالجينات .هذا الانزيم يقوم بتسريع الخطوة الاخيرة لتكسير حمض الاسكوربك (فيتامين سي ) .ان هذا الجين
مفقود في الانسان والرئيسيات وبعض المخلوقات الاخرى بما فيها الخفافيش ,
خنزير غينيا , وانواع معينة من الاسماك . بمقارنة هذا الجين الزائف بما
موجود في الجرذان فان النسخة البشرية وكذلك نسخة الالقردة الكبرى لديها
نقص فعال في الاكزونات في مناطق
I-III, V-VI, VIII, and XI وبمقارنة هذا مع الحذف المميز في الجين الزائف في خنزير غينيا والذي يشمل مناطق الاكزونات I, V, and VI
والتي تتطابق في نفس الطفرات .بالاضافة الى ذلك فان كودونات التوقف في
خنزير غينيا بتسلسلات 3TGA و 1TAA والتي معرفة في خنزير غينيا مشتركة
وفي نفس المواقع على الجين الزائف GULO في الرئيسيات بالطبع، يبدو أن
نحن البشر قادرون على ان نمضي على ما يرام دون هذا الجين لأننا أكل
الكثير من الأطعمة التي هي غنية في فيتامين سي مثل الحمضيات. فاذا كان
الامر كذلك ما هو الموضوع المهم ؟ حسنا المحاججة تسير بهذا الشكل (كما
يذكر ادوارد اي ماكس في مقالة في حديث الاصل )في معظم الثدييات يوجد جين
GLO فاعل موروث-بحسب نظرية التطور – من جين GLO الفاعل في السلف المشترك
.بوحسب هذه النظرة فان نسخ الجين الموجودة في الانسان وخنزير غينيا قد تم
تثبطها بواسطة طفرات .والتي يفترض انها حصلت بصورة منفصلة في كل من
الانسان وخنزير غينيا حيث كانت وجباتهم الطبيعية غنية بحمض الاسكوربك ولم
يكن غياب نشاط الانزيم GLO يمثل انعدام فائدة او يشكل ضغط انتقائي بالضد
من الجين المفقود .
علماء الوراثة الجزيئية الذين فحصوا الجين منو جهة نظر تطورية يعرفون
بان الطفرات الحذفية نادرة , لذلك فانهم كانوا يتوقعون وجود نسخ طفرية غير
فاعلة –جينات زائفة – كبقايا للجين الفاعل . وايضا فان نظرية التطور كانت
لتؤي الى التوقع القوي بان التغيرات الموجودة جين الانسان نفسها موجودة
في الرئيسيات (كالقردة والغوريلا ) . تم اجراء فحص لهذا التوقع واخذ مقطع
صغير من الجين الزائف GLO وتمت دراسته من كل من الانسان والاورانغوتان
والشيمبانزي والماكاكو .كل الجينات الزائفة الاربعة وجدت انها تمتلك طفرة
حذف لنيوكليوتيد مفرد مشتركة بينها تؤدي بباقي البروتين ان تتم ترجمته في
اطار قراءة ثلاثي خاطئ.
(Ohta and Nishikimi BBA 1472:408, 1999). 11,20
انه من المثير للاهتمام انه من بين طفرات التبديل العديدة المختلفة في
الجين الزائف GLO فان العديد منها –لكن ليس كلها – مشتركة وتشمل كذلك طفرة
حذف مفردة واحدة مشتركة بين كل الرئيسيات (عند مقارنتها بالجرذان ) .اذا
لم يكن السبب هو السلف المشترك فلماذا اذا تظهر تسلسلات الانسان
,الشيمبانزي , الاورانغوتان والوريلا طفرات حذف لنيكليوتيد مفرد في الموقع
97 من المنطقة المشفرة للاكسون اكس ؟ما هي الاحتمالات انه وبصورة عشوائية
من بين 165 زوج قاعدة تحدث الطفرة في نفس القاعدة ؟اليس هذا دليل لا ينقض
على وجود سلف مشترك ؟
هذه كانت الفكرة عند المقاربة الاولية .لكن نفس المجموعة البحثية اليابانية نشرت التسلسل الكامل للجين الزائف
GLO في عام 2003 م والذ من المفروض انه تطور بشكل عن الانسان وتمت مقارنته مع الجين الزائف في الانسان .
[Inai et al, 2003]
بصورة مفاجئة الفريق سجل العديد من الطفرات المشتركة (حذف وتبديل ) بين
الانسان وخنزير غينيا .ينبغي التذكر ان الانسان وخنزير غينيا انفصلا عن
بعضهما من السلف المشترك في نفس الوقت من انفصالهما مع القوارض .لذلك فان
الفروقات في الطفرات بين خنزير غينيا والجرذان لا ينبغي ان تشترك مع
الانسان باحتمالات اكثر عشوائية .لكن هذا لم يكن ما تمت ملاحظته , تمت
ملاحظة العديد من الطفرات المشتركة من بينها تلك التي في الموقع 97 . بحسب
ايناي ومجموعته فان هذا ضرب من الانحياز غير العشوائي غير المعتمد على
سلف مشترك اوسلف تطوري . احتمالية حصول طفرات التبديل في مواقع مختلفة في
الانسان وخنزير غينيا تمت حسابها من قبل ايناي ومجموعته لتكون 1,84*
10^-12 متماشية مع نقاط التطور الساخنة .
ما هو مثير للاهتمام هنا هو ان النقاط الساخنة الطفرية التي تمت
ملاحظتها بين الانسان وخنزير غينيا تتطابق مع تلك المجموعة التي تميز
الانسان والرئيسيات عن الجرذان .هذا المظهر المخصوص ابرز النقاش حول
القضية التي فهمها ايناي ومجموعته بصورة خاطئة .ريد كارترايت لاحظ عيبا
منهجيا في ورقة ايناي .
"على الرغم من ان الورقة البحثية لايناي ومجموعته فيها اخطاء منهجية
الان انهم فشلوا في الاخذ بعين الاعتبار ان التبديلات ممكن ان تحدث في
الجرذان بعد ان انفصلتمن الاثنين الاخرين . الباحثين في الواقع راكموا
تبديلات خاصة بالجرذان وقارنوها بتبديلات منفصلة مشتركة في خنزير غينيا
والانسان .
لو اجريت انا التحليل مكان ايناي ومجموعته لكنت استنتجت ان هنالك عشر
مواقع بين الانسان وخنزير غينيا حيث جرت تبديلات منفصلة لنفس النيوكليوتيد
, من ناحية اخرى تعرف بصفات مشتقة مشتركة .هذه المواقع هي 1 , 22 ,31 ,58
, 79 ,81 , 97 , 100 , 109 ,157 .مع ذلك معظم هذه تظهر لتكون تبديلات في
سلالة الجرذان عند النظر الى عينات اكبر من الانواع .
عندما ننظر الى جدول البيانات الواسع هذا , فقط موضع واحد من العشرة ,
81 , يظل صامدا كمثال على الصفات المشتقة المشتركة , 97 , غير حاسم , اما
الثمانية المتبقين فهي مواضع موروثة مشتركة . مع هذه المعلومات الاضافية
ااكون قد اظهرت ان "النقاط الساخنة " لايناي ومجموعته لعام 2003 م ليست
مدعومة بصورة جيدة "

انه يبدو بالفعل ان بعض التسلسلات التي لوحظن من قبل كارترايت هي فريدة
من نوعها في الجرذان – خصوصا اذا ما ضمنا المقارنة انواع اخرى . ومع ذلك
فان لي سؤالا في هذا الجانب . بانه هنالك وببساطة العديد من المواضع التي
يكون فيها تسلسل الجرذ هو الغريب الوحيد في الاكزون اكس (بمعنى اخر فان
هنالك سبعة او بصورة جدلية ثمانية مواضع اخرى ) .وباستخدام المعطيات
المنشورة عن معدل الطفرات والتي تقدر ب 2*10 ^-10 (درايك ) للموضع في
الجيل كنا لنتوقع رؤية طفرة او طفرتين من بين الاكسزون ذو ال 164
نيوكليوتيد موضوع المسالة (الاكزون اكس ) على المدة المفترضة المقدرة ب 30
مليون سنة .لذلك فان الافتراض بان اختلاف الطفرات في الجرذ يقترح وجود
معدل طفرات عالية في الجرذ عنه في خنزير غينيا .نفس الشئ يكون صحيح اذا
قارنا الجرذ بالفار (بمعنى ان معدل الطفرات هو اعلى في الجرذ منه في الفار
) .
هذا مثير للاهتمام بصورة خاصة حيث ان الكثير من الطفرات هي مرادفة
زلماذا بصورة اساسية بعض الطفرات تكون ثابتة في حوض جينات الجرذ اذا ما
قورنت بحوض جينات انواع اخرى .الا يعني معدل الطفرات العالي في الجرذ وجود
منطقة طفرات ساخنة ؟على الاقل في الجرذ .
بعيدا عن هذا فان الكثير من الطفرات هي ليست حصرية في حوض جينات الجرذ /
الفار وبالتالي فانها تقترح نقاط طفرات ساخنة بعيدة عن السخونة العامة
الكلية او ممكن ان تكون ميول للطفرات في هذا التسلسل الجيني المخصوص .
البعض لاحظ انه على الرغم من الطفرات المشتركة التي نشات بفعل النقاط
الساخنة للطفرات فانه لا يزال هنالك العديد من الاختلافات الطفرية بين
الانسان وخنزير غينيا / الجرذ مقارنة مع القردة . لذلك وبغض النظر عن
النقاط الساخنة فانه لا يزال الانسان والقردة اقرب الى بعضهما من منهما
الى الجرذ/ خنزير غينيا .
المشكلة مه هذه الحجة هو انه معدل الطفرات مرتبط مع المعدل العام لزمن
الجيل .تلك الكائنات التي لديها زمن جيل اقل لديها معدل طفرات اعلى خلال
نفس الفترة المجردة من الزمن مثلا 100 سنة . للذلك فانه من المتوقع ان
الكائنات بزمن جيل اطول نسبيا تتمتع بفروقات تطورية قليلة مقارنة مع
الكائنات الاقصر عمرا كالجرذان وخنزير غينيا . ما هو مثير للاهتمام حول
هذه الخسائر الطفرية انها تشترك بنفس التغيرات الطفرية .على الاقل انه
ظاهريا من المعقول القول بان طفرات الجين الزائف GULO
نتجت من عدم الاستقرار الجيني المتشابه المشترك بين بعض الكائنات
(كالانسان والقردة ) .نفس نوع الاشياء يمكن مشاهدته في لدرجة كافية في
منطقة GULOالعديد من الطفرات المناطقية مشتركة ما بين الانسان وخنزير
غينيا .اخذا بعين الاعتبار هذا العرض مرة ثانية :
لماذا يتشارك البشر وخنازير غينيا طفرات حذف كبيرة في الاكزونات
I ,V ,و VI بالاضافة الى اربع كودونات توقف لو كانت هذه الطفرات فعلا
عشوائية ؟بالاضافة لهذا فان مجموعة من الخنازير الدنماركية اظهرت ايضا ضياع
لوظيفة الجين الزائف GULO وخمنوا ماذا ؟ المفتاح لهذه الطفرة كان ضياع
جزء من الاكسزون الثامن هذا الجزء يطابق الجزء المفقود من الاكزون الثامن
للرئيسات .بالاضافة الى انه هنالك نقل اطاري للانترون الثامن ادى الى خلل
في تشفير الاكزونات 9 – 12 هذا يعكس خسارة مشابهة جدا في هذه المنطقة
للرئيسيات .هذه المفاتيح المتشابهة بصورة واضحة لم تنتج من سلف مشترك .هذا
يبدو انه دليل على ان العديد من الطفرات ان تكن جميعها هي نتيجة لمنطقة
GULO هي بالفعل نتيجة عدم استقرار جيني والذي جعلها عرضة لطفرات مختلفة
بالاخص في الانواع المتشابهة .
كشئ جانبي . العديد من الطفرات الجينية والتي تنتج خسارة وظيفية معروف
انها تصيب نفس المواضع الجينية بنفس الاسلوب بعيدا عن مسالة السلف المشترك
.على سبيل المثال مرض اكوندروبلاشيا (نقص الغضاريف ) يحصل نتيجة طفرة
تلقائية في الانسان بنسبة 85 % من الحالات .في الانسان يحصل المرض نتيجة
طفرة في الجين FGFR2
ملاحظة مهمة في هذا الجين هو ان الطفرات تحصل في منطقتين هما
(755 C->G and 755-757 CGC->TCT)
بصورة مستقلة عن السلف المشترك .الساق القصيرة للكلب الالماني حصلت بطفرات مشابهة كما ان طفرات مشابهة حصلت للخرفان .
Real Time Molecular Convergence
مثال
آخر للاهتمام لهذه الظاهرة تمت دراسته بالتفصيل في الكائنات الاسرع
تكاثرا، مثل الفيروسات. على سبيل المثال، نشرت دراسة مثيرة للاهتمام من قبل
بول وآخرون، عن خط الأنساب المضاعف للفيروس الملتهم للبكتيريا phiX174.
وقعت العديد من الطفرات في كل جينوم خلال الانتشار . حيث انه عبر تسعة
أنساب منفصلة وقع 119 بديل مستقل في 68 موقع للنوكليوتيدات. ما يثير
الاهتمام هنا هو أن أكثر من نصف هذه التبديلات في ال1/3 من هذه المواقع
كانت متطابقة في الأنساب المختلفة. وكانت بعض التبديلات المتقاربة محددة
إلى مضيفين محددين في حين أن تبديلات اخرى كانت مشتركة بين مضيفيان
منفصلان. ااعادة التركيب للعلاقات بين الانواع باستخدام تسلسل الجينوم
الكامل لم يفشل فقط في استعادة التاريخ التطوري الصحيح بسبب هذه التغييرات
المتقاربة، ولكن تم رفض التاريخ الحقيقي على أنه أدنى إلى حد كبير ليتناسب
مع البيانات. في دراسة لاحقة مماثلة بول وآخرون يقولون إن مثل هذه النتائج
"تشير إلى عدد محدود من المسارات التي اتخذت خلال التطور في هذه
الفيروسات، وتثير أيضا احتمال أن الكثير من التباين في الأحماض الأمينية في
التطور الطبيعي لهذه الفيروسات كانت انتقائيا". 29 وبعبارة أخرى، فإن
الكثير من الاختلافات في العوامل الوراثية الفيروسية ليست محايدة، بل هي في
الواقع وظيفية وتم الحفاظ عليها عن طريق الانتقاء الطبيعي.
هذا مدهش!
الآثار هنا مذهلة للغاية. إذا كانت الطبيعة المتقاربة للطفرات الجزيئية
مثل هذه لا يمكن الكشف عنها بصورة كافية فانها ستتداخل مع أي شكل من اشكال
رسم شجرة العلاقة بين الانواع يعتد بها أو تحديد دقيق للعلاقات التطورية.
إذا كان هناك أي نوع من العلاقة مع الكائنات متعددة الخلايا الاعلى مستوى ،
فان هذا يمكن ان يقوض علم الاحياء التطورية باكمله كما هو متعارف عليه
اليوم . وهناك في الوقت الحالي حاجة إلى دراسات مثل هذه على نطاق واسع
لتحديد ما إذا كانت الطفرات المتقاربة مثل هذه أكثر انتشارا. ومن الواضح،
فإن الافتراض الشائع أن الطفرات المتقاربة على المستوى الجزيئي نادرة
ونتيجة لفرصة عشوائي تماما هو ببساطة ليس صحيحا بعد الآن على الاقل في بعض
(وربما معظمها إن لم يكن الكل) العوامل الوراثية.
نتائج مماثلة تم
الكشف عنها في الآونة الأخيرة من قبل كويفاس وآخرون. في عام 2002 نشر مقالة
في مجلة علم الوراثة Genetics تتكلم عن فيروسات RNA (انظر الملحق) . في
هذه الدراسة الكتاب أثبتوا مرة أخرى أ التقارب في 12 موضع مختلف في الأنساب
المستقلة. فوجئ الكتاب باكتشاف أن التقارب حدث ليس فقط داخل المواضع غير
المترادفة، ولكن في مواضع مترادفة ومناطق داخل الجين intergenetic كذلك
(كان يعتقد عادة أنها محايدة بالنسبة لكونها من الانتقاء الطبيعي) كما لاحظ
القائمون أن لا يقتصر هذه الظاهرة إلى المختبر، ولكن أيضا على تم ملاحظته
على نطاق واسع نسبيا في فيروس HIV-1 استنساخ الفيروس في البشر وفي سلالات
معزولة (SHIV) من قرود المكاك شيف، والقرود، والبشر.
نفس المؤلفين
مضوا قدما ليلاحظوا أن "التطور المتقارب على المستوى الجزيئي ليس مثيرا
للتناقض طالما يمكن التوفيق بين ذلك معالنظريات المحايدية والنظريات
الانتقائية . النظرية المحايدية تشير إلى أن التقاربات هي ببساطة حوادث، في
حين في إطار الانتقائية ، فان هنالك مؤهلين للتقاربات. التفسير الأول يرى
التقارب بأنه تكيف وكنتيجة لكون الكائنات المختلفة تواجه نفس البيئة (كما
في حالة تجاربنا) مع مسارات بديلة قليلة من التكيف (كما هو متوقع للجينوم
المضغوط). الثاني، مع الأخذ في الاعتبار نموذج التدخل نسيلي، والطفرات
المفيدة يجب أن تصبح مثبتة بطريقة منظمة (Gerrish وLenski 1998)، حيث ان
افضل مرشح ممكن يثبت أولا، ثم ثاني أفضل مرشح، وهلم جرا، وهذا يفرض، انه
باعطاء حجم سكان كبير بما يكفي لجعل التداخل النسيلي عاملا تطوريا هاما،
ينبغي لنا أن نتوقع دائما أن تكون نفس الطفرات ثابتة. "
وفقا للكتاب،
فان الحجة السابقة هي صالحة لتغييرات غير المترادفة ولكن يجب إيجاد تفسير
بديل للتغييرات المترادفة او التغيرات في المناطق بين الجينات وبما أن هذه
التغيرات عادة ما تكون محايدة بشكل انتقائي. لذا، فإن الكتاب يشير إلى أنه
"RNA الجينوم والذي يشارك في العديد من تفاعلات RNA - RNA و RNA
-البروتين التي تؤثر على تكاثر الفيروس. وهذا واضح للمناطق، غير المشفرة
المنظمة (ستيلمان و ويت 1997، 1998)، ولكن هناك أدلة متزايدة على أن مناطق
التشفير القفيصة في فيروس بكورنا picornaviruses قد يكون لها أيضا تأثير
على تكاثر الفيروس (ماكنايت و ليمن 1998؛ فيرس وآخرون 2001). لذلك، قد يكون
ال RNA نفسه (وبصرف النظر عن القدرة تشفير البروتين التي يمتلكها) يساهم
في اعطاء النمط الظاهري الفيروسي، واللياقة ويمكن أيضا أن تتأثر
بالتبديلات المترادفة. " هذه نقطة مهمة لأنه "قد تم الاعثور على ادلة
لانتقاء المواضع المترادفة ايضا في الثدييات (اير ووكر 1999)، نتيجة
للانتقاء الذي يؤثر على المكونات الاساسية لمخطط الحرارة والضغط وأجزاء
كبيرة من الحمض النووي الخردة."
وبعبارة أخرى، لا يبدو أن الكثير DNA،
حتى في المناطق التي تبدو غير وظيفية من الحمض النووي أو حتى بين التغييرات
المترادفة، وهو حقا غير وظيفي عندما يتعلق الأمر بالجينوم الفيروسي.
الكتاب اذا انتقلوا إلى اقتراح المقارنة مع الجينوم من كائن رفيع المستوى،
مثل شبيه الانسان.
"على سبيل المثال، ذكرت فاي وآخرون (2001) أنه في
البشر، فإن الغالبية العظمى (80٪) من التغيرات الحمضية الأمينية هي ضارة
إلى حد ما وفقط جزء صغير محايدة. ومن بين هذه الطفرات الضارة للاحماضي
الأمينية، ما لا يقل عن 20 ٪ مؤذية بشكل طفيف . هنا، وجدنا أن 15 موضعا
للأحماض الأمينية يتغير، مع كونها مفيدة فقط في 5 مواضع بشكل كبير. عند هذه
النقطة، يمكننا التكهن فقط حول الدور الانتقائي لجميع مواقع الأحماض
الأمينية والتي تبين أنها غير متغيرة في دراستنا. و إجمالي عدد الأحماض
الأمينية في خمس جينات من VSV هو 3536 على افتراض أن التغيرات في أي من
3536 -. هو 15 فان 3521 الأحماض الأمينية الثوابت سيكون ضارة (وبالتالي تم
ابعادها عن طريقالانتقاء التنقوي خلال التجربة أو التطور)، ومن ثم نسبة
تبديلات الاحماض الأمينية التي يمكن أن تكون ضارة يكون 3521/3536 = 99.58٪
~؛ فإن كسر المواقع المحايدة يكون 10/3536 = 0.28٪ ~؛ بينما فقط 5/3536 =
0.14٪ ~ سيكون مفيدا على الرغم من الاختلافات بين البشر و VSV في حجم
الجينوم وتنظيمه وطبيعة الحمض النووي المستخدمة، في كلتا الحالتين فإن نسبة
البدائل الضارة المحتملة للأحماض الأمينية أكبر من ذلك بأغلبية ساحقة من
تلك المحايدة أو المفيدة. "
وبعبارة أخرى، فإنه على الأقل معقولة
الاشتباه في أن القليل جدا من الحمض النووي المشفر ، وحتى في الهومينيد، هو
حقا "محايدة" أو في مأمن من كل ضغوط الانتقاء الطبيعي. هذا اصبح صحيحا حتى
من ناحية الDNA غير المشفر الذي كان يعتقد انه خردة وجدت لتكون وظيفية
(لينك). هذا يوحي بقوة بأن العديد من ما كان يعتقد أن تكون أخطاء طفرية
مشتركة قد يكون في الواقع تم الحفاظ عليها وظيفيا، تحتفظ بها مخلوقات
مماثلة في بيئات مماثلة. في ضوء ذلك، والنظر في الاستنتاجات التالية من وود
وآخرون نشرت في طبعة عام 2005 من مجلة Genetica:
الدليل الأكثر إقناعا
على التكيف الجيني الموازي يأتي من التجارب التي تنطوي على انتقاء اصطناعي
يتضمن مجموعة سكان ميكروبية. في بعض التجارب، حوالي نصف بدائل
النوكليوتيدات الموجودة في الأنساب مستقلة تحت انتقاء منتظم هي نفسها.
دراسات العلاقة بين الانواع توفر وسيلة لدراسة التكيف الوراثي الموازي في
الانظمة غير التجريبية، ولكن دليل قاطع قد يكون من الصعب الحصول عليه لان
الhomoplasy (نتائج التطور التقاربي )يمكن أن تنشأ لأسباب أخرى. ومع ذلك،
قدمت المقاربات من دراسة العلاقة بين الانواع دليلا على التكيف الوراثي
الموازي في جميع المستويات التصنيفية، وليس فقط الميكروبات. المقاربات
الكمية الجينية تشير أيضا إلى التطور الوراثي الموازي عبر كل الأصناف سواء
القريبة و البعيدة الصلة، ولكن من المهم أن نلاحظ أن هذا النهج لا يمكنه
التمييز بين التغيرات في مواضع متوازية متماثلة مقابل التغييرات في مواضع
متقاربة غير المتماثلة ولكنها ترتبط ارتباطا وثيقا. والنتيجة أن التكيف
الوراثي الموازي يبدو أنه يحدث بشكل متكرر وعلى جميع المستويات التصنيفية
له آثار هامة بالنسبة للدراسات التطورية ودراسات العلاقة بين الانواع
.بالاخذ بالاعتبار التحليلات فيما يتعلق النشوء والتطور، فان التغيرات
الوراثية المتوازيةلو كانت شائعة، يؤدي إلى تقديرات خاطئة عن علاقات النشوء
والتطور. [التشديد مضاف]
لاحظ أنه وفقا لوود وآخرون، فان الطفرات
المتوازية مع / أو الطفرات المتقاربة هي "متكررة" في "كل المستويات
التصنيفية، لا مجرد الميكروبات". هذا مثير جدا للاهتمام ولديه بالفعل آثار
خطيرة جدا عندما يتعلق الأمر بتحديد علاقات النشوء والتطور مع بعضها التي
من المحتمل أن تكون ليست فقط خاطئة، ولكن لا معنى لها بقدر ما نظرية النشوء
والارتقاء من أصل مشترك معنية. بدلا من ذلك، قد يكون أكثر أوجه التشابه من
ناحية النشوء والتطور انعكاس لأوجه التشابه والاختلاف الوظيفية منها
انعكاسا صحيحا لعلاقات تطورية .
النقاط الساخنة الطفرية
بالعودة
إلى النقاط الساخنة الطفرية،نتسال ما الذي يجعل تلك النقاط الساخنة
"ساخنة"؟ لعل الجواب يكمن في الطبيعة الكيميائية للمنطقة الساخنة. و نوع
الروابط الجزيئية، واستقرارها، أو عدم استقرارها، أو غيرها من التفاعلات
الجزيئية تصلح لمفاتيح معينة لازاوج النوكليوتيدات، وتغيرات بيئية معينة
معطاة بصورة خاصة . لا أحد يعرف على وجه اليقين حقا إلا أن يقول إن النقاط
الساخنة الطفرية موجودة بالفعل. لذلك، باعتبار أنها موجودة بالفعل، ينبغي
أن يتوقع ان الجينات المتشابهة تعمل بطرق مشابهة، وهذا يشمل وجود مماثلة
طفرية "النقاط الساخنة و / أو" أخطاء مشتركة. " وعلى أية حال، فإنه من
المثير للاهتمام أن نلاحظ أنه لا توجد مثل هذه الأمثلة من "الأخطاء
المشتركة" بين الثدييات ومجموعات أخرى من الحيوانات (على الرغم من أن هناك
الكثير من "الأخطاء" المشتركة التي يتم مشاركتها من قبل جماعات متباعدة على
نطاق واسع في الثدييات).
لا توجد أمثلة على 'أخطاء مشتركة' تربط
الثدييات إلى فروع أخرى من شجرة النسب الجيني للحياة على الأرض. . . ولذلك،
يجب على العلاقات التطورية بين الفروع البعيدة على شجرة النسب الجيني
التطورية genealogic ان تقوم على أدلة أخرى إلى جانب 'الأخطاء المشتركة. "
طبعا
الحجة المستخدمة لشرح هذه الحقيقة هي أن الثدييات انفصلت من مجموعات أخرى
من الحيوانات منذ أكثر من 200 مليون سنة مضت. ونظرا لهذا المقدار من الوقت،
فان الطفرات العشوائية طمست اي اثر للأخطاء الجينية الشائعة. هذه نقطة
جيدة جدا. لكن يبقى السؤال لماذا يتم يتم الاحتفاظ ببعض الأخطاء الجينية
المحددة طالما أنها في الواقع عديمة الوظيفة؟ أيضا، "الجينات الزائفة
المعالجة" مشابهة جدا "للعناصر الجينية المنقولة" التي تنتقل غالبا من
حيوان الى اخر عن طريق الفيروسات . بعض الجينات الزائفة داخل الانواع قد
شارك في الحقيقة سلف مشترك بينما الانواع المختلفة من الحيوانات نفسها،
التي تؤوي بعض هذه التسلسلات الجينية، ربما تكون غير مرتبطة من خلال أصل
مشترك بقدر ما ترتبط جزئيا من خلال أنها عدوى شائعة.
وعلى أية حال، فانه
لا يوجد في الواقع علامات وراثية بصورة سهلة جدا "foolproof"لاصل مشترك.
وقد تبين أن جميع العلامات السهلة المقترحة حتى الآن فيها عيوب كبيرة.
التنبؤ بأن الجينات الزائفة، ترانسبوزونات (SINEs و LINEs) وغيرها من
الأخطاء الطفرية المشتركة هي دليل قاطع على الأصل المشترك لم يعد مقبولا
على مدى السنوات الأخيرة. على سبيل المثال، النظر في المقتطف التالي من
ورقة ديفيد حلس "بعنوان" "SINEs ذات الشخصية المثالية ". نشرت في دورية
الاكاديمية الوطنية للعلوم، 1999:
ماذا عن الادعاء بأن أحداث الإدخال
SINE/LINEهي علامات مثالية للتطور (أي أنها لا تظهر homoplasy)؟ وقد حصلت
ادعاءات مماثلة لأنواع أخرى من البيانات في الماضي، وفي كل حالة تم العثور
على أمثلة لدحض هذا الادعاء. على سبيل المثال، كانت البيانات تهجين ال
DNA-DNA ;HK كان من المفترض أن تكون في فترة ما في مأمن من التقارب، ولكن
تم اكتشاف العديد من مصادر التقارب لهذه التقنية. ويعتقد أن إعادة الترتيب
الهيكلية للجينوم كان يفترض ان تكون احداثا معقدة بحيث ان التقارب كان من
غير المرجح للغاية، ولكن تم اكتشاف العديد من الأمثلة الآن على التقارب في
إعادة ترتيب الجينوم. وحتى احداث الحذف والادخال البسيطة داخل مناطق
التشفير اعتبرت من غير المرجح أن تكون مثلي الاستعاضة، ولكن من المعروف
الآن أمثلة عديدة من التقارب والتوازي من هذه الأحداث. على الرغم من ان
النيوكليوتيدات المفردة والاحماض الامينية معروفة على نطاق واسع بكونها
تظهر homoplasy،فان بعض الكتاب اقترح أن التقارب في وقت واحد على نطاق
واسع في العديد من النيوكليوتيدات يكاد يكون من المستحيل. ومع ذلك، فقد ثبت
وجودأمثلة على مثل هذا التقارب في الدراسات التجريبية للتطور .
فيروس النسخ العكسي الداخلي ERV
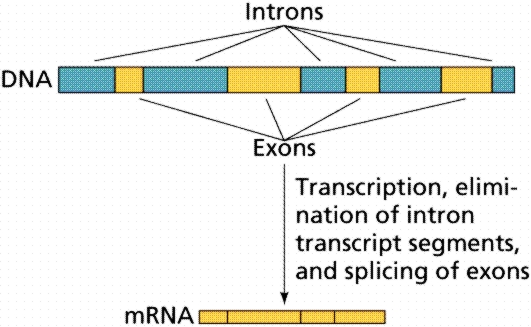
مصطلح "إنترون" هي اختصار ل "المنطقة داخل الجين"، والتي هي أجزاء من
الحمض النووي DNA داخل تركيب ال DNA ترمز او تشفر للجينات. الإنترونات
شائعة في الكائنات حقيقية النواة (الحيوانات والنباتات والفطريات، الخ)،
ولكن ليس في بدائيات النوى (البكتيريا وغيرها) عندما يتم نسخ الجينات
المحتوية على الإنترونات، الإنترونات عادة تقسم، وتاركتا نسخة من تسلسل
الحمض النووي دون تسلسل إنترون الأصلي. حجم الإنترونات يتارجح على مدى واسع
جدا، من اقل من 20 زوج قاعدة الى حوالي 500،000 زوج قاعدة. الأمر اللافت
أيضا هو أن الحجم الإجمالي للالإنترونات ضمن الجينات نظرا قد يكون أكبر
بكثير من المناطق المشفرة للجين نفسه - أكثر من 90٪ في بعض الأحيان.
الأمر
المثير لاهتمام أيضا حول الإنترونات، لأغراض هذه المناقشة وجه الخصوص، هو
أنه لفترة طويلة كان يعتقد أن الإنترونات بقايا تطورية من الادخالات
العشوائية للحمض النووي -الادخالات العشوائية التي هي دائما تقريبا إما
ضارة أو محايدة. بطبيعة الحال، إذا كان الإدخال محايدا أو قرب المحايد،
يمكن أن يصبح ثابتا في جينات السكان وأن تنتقل مع مرور الوقت. وبشكل عام،
ومع ذلك، بسبب فكرة أن إدخال إنترون هي دائما تقريبا ضارة، تم افتراض أن
أحداث الإدخال مثل هذه نادرة للغاية.
ومع ذلك، فان بعض البحوث
المثيرة للاهتمام من قبل لي وآخرون . وواللتي نشرت في عام 2009 اشارت إلى
أن ادخالات الإنترون ليست نادرة، أو تضر عادة، أو نتيجة لأصل مشترك اذاما
وجدت في نفس الموقع داخل الكائنات الحية المختلفة.
" التحليلات
الجزيئية المتوفرة لدينا تدفعنا لرفض العديد من الفرضيات حول آلية أصول
الإنترون، في حين تشير بوضوح الى مسار غير متوقع تماما - الظهور كحوادث
بزغت اثناء اصلاح التكسرات في الامتداد المزدوجللحمض النووي"
واضعو
هذه الدراسة لاحظوا أيضا أن 17٪ من عمليات الإدخال في الإنترونات المرصودة
كانت الإدخال يتم بصورة متوازية متطابقا في الانواع الجينية المستقلة او
خطوط الانساب المستقلة , بعبارة اخرى فان ادخالات الانترون تفضل مناطق
ساخنة معينة في الجينوم
"اكثر النتائج غرابة بالنسبة لي هي الحالات
المتعددة للكسب المتوازي للانترون، لأن هذا يعني أن الغار (Daphnia) هو في
طور نشط من تكاثر الإنترون،" كما يقول لى. "وهذا يجعل الDaphnia نظام فوق
العادي لدراسة تطور الإنترون. وبالإضافة إلى ذلك، فإننا نعتقد بان عملنا
يسهل على تقدير أكثر دقة لمعدلات كسب الإنترون، ويتحدى بصورة مباشرة
الافتراض بأن مكاسب الانترون الموازية هي نادرة كما تقول العديد من
التحليلات السابقة ....
بشكل ملحوظ، وجدنا العديد من حالات مكاسب موازية
في مواقع إنترون نفسها بصورة اساسية في انواع جينية مستقلة، "وقال لينش".
هذا يحتج بشدة ضد الافتراض الشائع أنه عندما يشترك نوعين مختلفين بنفس
الإنترونات في نفس الموقع،فان السبب هو دائما الميراث من سلف مشترك. "
القضية كدليل على السلف المشترك
الفيروسات النسخ العكسي داخلية المنشأ أو "ERV" عناصر الفيروسية التي
يعتقد أن أدرجت نفسها في الجينوم من المخلوقات المختلفة - لتشمل البشر
والقردة. ويعتقد العديد من المراقبين أن ERV من بين أقوى الأدلة التي تدعم
نظرية الأصل المشترك. على سبيل المثال، ثمة من يقول أن أفضل تفسير لوجود
نفس الفايروس في نفس المواقع في الجينوم في كل من البشر والقردة هو ان
كلاهما انحدر من أصل مشترك المشتركة بين البشر والقردة. وبعبارة أخرى، يجب
أن يكون قد سلف مشترك من البشر والقردة واحد لإدراج ERV في الجينوم. ثم،
في وقت لاحق، عندما أسلاف البشر والقردة العليا انشقت من سلالة الأسلاف
المشتركة هذه، حافظ تسلسل ERV نفسه في نفس الأماكن في كل من الجينوم
للذرية.
هذه الحجة تبدو واضحة ومباشرة عند المقاربة الاولية. ومع ذلك، فان هناك العديد من المشاكل المحتملة مع هذه النظرية.
العديد من المشاكل المحتملة مع هذه النظرية.
علامات الوظيفة
واحدة من المشاكل هو ان بعض فيروسات النسخ العكسي او على الاقل اجزاء من فايروس النسخ العكسي اكتشف انها لها فائدة وظيفية .
فايروس النسخ العكسي كما في enJSRV وجد انه يقوم بتنظيم نمو وتمايز
الاديم الظاهر الغاذي في ما قبل الحمل عند الغنم . هذا العمل دعم الفرضية
بان فايروس النسخ العكسي يلعب دورا في تشكل المشيمة وتكاثر الثدييات "
في الحقيقة فان عناصر فيروس النسخ العكسي يعتقد انهاتتحكم او تساعد في
في التعبير عن 20 % من جينوم الانسان .وممكن ان تحفز الانتهاء المبكر
للتعبير على مسافة .انه ايضا من المثير للاهتمام ان
ERV وعناصر حمض نووي اخرى طفيلية مفترضة موجودة غالبا في جينوم
الكائنات الاكثر تعقيدا تقترح مجددا بان المناطق غير المشفرة من الحمض
النووي والتي اعتقد سابقا انها بقايا تطورية "خردة " هي في الواقع تلعب
دورا وظيفيا في جينوم الكائنات المعقدة وظيفيا .
"مع تراكم بيانات تسلسل الجينوم فان بعض طرز تطور الجينوم غير المشروحة
بدات بالظهور. من الملاحظات الصادمة هو الميل العام للكائنات العليا
لتطوير كثافة جين قليلة مع الرتبة الاعلى . على سبيل المثال بكتيريا
E. Coli لديها كثفاة جين ب 2 كيلو قاعدة للجين . Drosophila
4 كيلو قاعدة للجين . الثدييات 30 كيلو قاعدة للجين . الكثير من نقصان
الكثافة سببه كثرة تراكم الحمض النووي غير المشفر او الطفيلي كما في النوع
واحد او اثنان من الترانزبوسون .النظرية التطورية الحالية لا تدعم بصورة
كافية هذه المشاهدات . بالاضافة الى ذلك فان الثدييات يبدو انها احتفظت
بعلى الاقل بعض النسخ غير المنقوصة "الجينوم "من الفايروس العكسي .كما في
الدقائق نوع اي او ERV انه من السائد الان انه لا يعرف الضغط التطوري الذي
ادى الى الاحتفاظ بهذه الفيروسات الجينومية.
على نفس الخط ورقة لاحقة قدمت الدليل الذي يقترح الاسباب لذلك "الضط الانتقائي " الذي تمت ملاحظته والحاجة الفعلية ل ERV
بداخل جينوم الكائنات المعقدة كالانسان .الورقة عرضت في مجلة المعلومات الحياتية Bioinformatics
كتب Conley et. Al.
" لقد رصدنا 51,197 تسلسلات بادئةمشتقة من فيروس العكسي تبدا التعبير
في جينوم الانسان من بينها 1743 حالة حيث يبدا التعبير من من تسلسلات
فيروس العكسي تقع في البادئ القريب للجين او او المنطقة غير المترجمة 5
(UTRs) ان تحليلنا كشف بان تسلسلات الفيروس العكسي في الانسان تشفر لعشرات
الالاف من المحفزات النشطة الصور طبق الاصل للفيروس العكسي تناظر حوالي
1,16% من تسلسل جينوم الانسان وبطاقات PET التي تلتقط القطع المستنسخة طبق
الاصل والتي بدات من الفيروس العكسي تغطي 22,4 %من الجينوم .هذه البيانات
تقترح ان الفيروس العكسي ينظم الاستنساخ لدى البشر بدرجة عالية . وفيروس
النسخ العكسي الداخلي ERV وجد انه يحمي البشر من العدوى الضارة وبالفيروس
الخارجي exogenous "ان دورا احيائيا مفترضا ل
ERV هو انها تساعد المضيف على ان يقاوم العدوى المرضية للنسخة الخارجية
من الفيروس , معطيا فائدة منتقاة للمضيف الذي يحملهم .على سبيل المثال
بعض فيروسات النسخ العكسي الداخلي في الطيور والفئران ممكن ان توقف العدوى
بالفيروس العكسي الخارجي في مرحلة الدخول عن طريق تداخلها مع المستلمات .في الفئران فان Fv-1
يمنع العدوى في خطوة ما قبل التكامل ايضا ممكن رؤيته كفيروس نسخ عكسي"
فيروسات النسخ العكسي ممكن ان تساعد في تعيير نشاط النظام المناعي
"على
سبيل المثال تسلسل HERV-K لسرطان الخصية للانسان الفيروس المشتق منه نوع
(HTDV) تم تسجيل انه ممكن ان ينتج دقائق شبيهة بفيروس النسخ العكسي وممكن
ان ينتج جينات gag, pol , env
عن طريق دوال .ايضا فيروس النسخ العكسي 3 ممكن ان ينتج جين نوع
env
في الخلايا الجنينية للمشيمة .هكذا تقارير ممكن ان تشرح المشاهدات المبكرة
المتعددة لوجود دقائق فيروسية في انسجة الانسان .على الرغم من ان البعض من
HERVs ممكن التعبير عنها في اورام الام .فان الاورام السنورية RD114,
ERV-3, HERV K10+
جميعها يعبر عنها في النسيج المشيمي .ما هو المميز اذا
في الفيروس العكسي غير المنقوص ولماذا تعبيرها شائع هكذا في الاجنة ؟انا
وفينابلز ومجموعته من مجموعة بويد اقترحنا ان البعض من تلك ال HERV
ممكن ان تعمل خلال غرس الجنين لمنع التعرف على الجنين كعدو مناعي من قبل النظام المناعي للام .
بالاضافة
فان الجين gag لفيروس النسخ العكسي ممكن ان تكون منظمة للمناعة .حيث ان
p(70) gag الفئران تم نسخه وانتاجه ووجد انه يماثل عامل الالتصاق IgE
(IgE-BF)
والذي هو منظم للخلايا المناعيا نوع بي لانتاج الايميونوغلوبيولين نوع H
ومؤخرا جدا تم تسجيل ان ال gag الداخلي Fv-1 وهو شبيه Herv.L
فيروس النسخ العكسي الداخلي يحمل مقاومة للاورام MLV
وعلى الرغم من العديد من الباحثين يعارضون الدور المناعي لل
p15E
الا ان نشاط تثبيط مناعي في عينات الزرع قد تم رصده بصورة واضحة .هذه
النتائج تبدو كافية للتنبيه للقيام بالمزيد من التحقيقات للتاكد من ان
الجينات الناتجة من فيروس النسخ العكسي نوعي env و gag لها دور في تنظيم
المناعة
اذا
انه ليس بالضرورة انه كل التسلسلات الشبيهة بفيروس النسخ العكسي هي بقايا
تطورية غير فاعلة لعدوى فيروسية عشوائية كما يشيع ذلك التطورييين المشهورين
كريتشارد داوكينز واو دغلاس ثيوبالد . هذه الحقيقة تم تسليط الضوء عليها
من قبل ريشارد ستيرنبيرغ عام 2002 م في مجلة
Annals of the New York Academy of Sciences
في الجملة التالية
"ان
قصة الحمض النووي الاناني وما يؤيدها من اطر ينبغي ان تضم الى الايقونات
الاخرى لنطرية التطور للدارونيين الجدد والتي على الرغم من تباينها مع
الادلة التجريبيةعليها الا انها لا تزال موجودة في الكتب "
دعم مشابه لهذا المفهوم اورده د.وانغ من مختبر هاوسلر
"هذه
النتائج تطرح التساؤل حول دور "الحمض النووي الخردة " تلك المنطقة الواسعة
من الجينوم التي لا تشفر للبروتينات .ان فيروسات النسخ العكسي الداخلية
تقع ضمن التصنيف الذي لطالما اعتبرها العلماء كحمض نووي لا يخدم اي غرض
ولكن بينات جديدة من مختبر هاوسلر ومختبرات اخرى تتحدى هذه النظرة " .
هل اصل فيروسات النسخ العكسي الداخلية من الفيروسات الخارجية ام العكس ؟
هناك ادلة على ان الفيروسات الخارجية مشتقة من فيروسات النسخ العكس الداخلية بدلا من الطريق الاخرى .
"فيروسات النسخ العكسي الخارجية مشتقة من ERVs وعلى الخصوص
ERV-Ls تمثل وسيط بين الترانزبوسون العكسي والفيروس الخارجي.".
هذا
المفهوم مدعوم بالملاحظة انه لا توجد امثلة معروفة على وجود ادخالات جديدة
لفيروس خارجي في الانسان المعاصر.ايضا لا يوجد نظير خارجي معدي موجود
اليوم لفيروس النسخ العكسي الداخلي في الانسان . هذه نتيجة مثيرة للفضول في
ظل شوع وجود فايروس النسخ العكسي في جينومي الانسان والقرد فيما اذا كانت
الفرضية المنتشرة حول كون تسلسل فيروس النسخ العكسي الداخليفي الحقيقة مشتق
من الفيروس المعدي الخارجي
"لا يوجد في الوقت الحالي نشاط نسخي لHERVs او تحويل الى عنصر داخلي لفيروس نسخ عكسي خارجي للانسان قد تمت توثيقها منذ مدة "
"معظمها
يمثل عدوى فايروسية قديمة كما مدلل عليها بالانتشار الواسع في نوع
الرئيسيات ,ولا يوجد نظير معدي لفيروس النسخ العكسي الداخلي في الانسان
HERVsموجود اليوم ".
هذا يفتح الباب على احتمالية ان كل فايروسات
النسخ العكسي الخارجية اشتقت من ERVs ليس بالضرورة ان اطريق الاخر في كون
ال ERVs مشتق بالاصل من عدوى بالفيروس الخارجي .بكلمة اخرى فان فان كل
التسلسلات المحتملة والتي تمثل فايروسات او عناصر فايروسية ممكن ان تكون
اشتقت بالاصل من تسلسلات جينية فاعلة والتي عانت منذ فترة من تغيرات
اضمحلالية وفقدان للتحكم نتج عنها خواص طفيلية متنوعة نراها في الفايروسات
اليوم .بالاضافة الى التاثير الضار الناتج عن فقدان التحكم كما نراه في
تسببها بالسرطانات والاورام .
ادخال فيروسات غير عشوائي
بعيدا عن هذا تبين ان ادخال فايروس
النسخ العكسي الداخلي ليس بصورة عشوائيةبالرغم من الايمان العام كما يعتقد
من التيار السائد للعلماء .
فايروس النسخ العكسي يظهر ميلا لمواقع معينة في الجينومات المتنوعة .
"على
لارغم من ان تكامل الفيروس العكسي ممكن ان يحصل خلال الجينوم فانه يوجد
نقاط ساخنة محلية للتكامل حيث هنالك تفضيل قوي لمواضع محددة على الاخرى
ممكن اظهاره عن طريق الاحصاءات .الابحاث عن فايروس الايدز وسرطان دم
الفئران اظهرت وجود ميل للتكامل مع مناطق معبرة لجينوم المضيف .في حالة
فايروس سرطان دم الفئران في مواقع قرب بداية التعبير .اسس هذا التفضيل غير
معروفة ولكن يعتقد انها ممكن ان تعكس تفاعل لمعقد ماقبل التكامل مع
بروتينات معينة او تسلسل حمض نووي معين او تراكيب مرتبطة بالتعبير ".
"لكن
على الرغم من مفهوم انتخاب فايروس النسخ العكسي لمواقع معينة هو السائد
حاليا الا انه عمليا كل مواقع الجينوم ممكن ان تمثل اهداف للتكامل ,مع ذلك
بتفضيلات متباينة .كان هنالك مناطق ساخنة معرفة تم استخدامها اكثر ب280 مرة
عما كان متوقعا حسابيا
الاحتمالات في ادخالات فايروس النسخ العكسي في الخلايا الجرثومية
من
اجل ان يحصل ادخال فايروس النسخ العكسي في نفس مواقع الجينوم في مختلف
التجمعات السكانية على اساس السلف المشترك واحد من امرين يجب ان يكون حصل
.اما افراد عديدين في نفس التجمع السكاني قد اصيبوا بالعدوى في نفس الوقت
والعدوى استقرت في نفس المواقع من الجينوم ( سيناريو غير واقعي ) او ان
هنالك مرحلة عنق زجاجة مر بها التجمع السكاني (كان يكون فرد واحد ) اصيب
بالعدوى وانتقلت لذريته بالتعاقب فاستلزم ذلك الجيل كله حتى يتم تثبيت
تسلسل الفايروس في الجينوم الخاص بكل افراد السكان .
باختصار فان حدثا
فايروسيا ممكن يكون حصل لكل الافراد في التجمع السكاني لكل تسلسل من فيروس
النسخ العكسي في الجينوم (المئات او الالاف منهم ) وكل فايروس نسخ عكسي
داخلي توجب عليه ان يحقن نفسه دون ان يؤثر على اللياقة التكاثرية للمضيف
...وكل هذا يج ان يحدث لمرات عديدة في مختلف الانواع .فرص حدوث هذا الشئ لا
يبدو انها هكذا محتملة .خصوصا اذا اخذنا بالاعتبار الاحتمالات العالية
للطفرات المتلفة في الانسان والقردة والفرصة الحقيقية ان تؤدي تلك الطفرات
المتلفة الى الذوبان الجيني والانقراض مع توسع السكان .
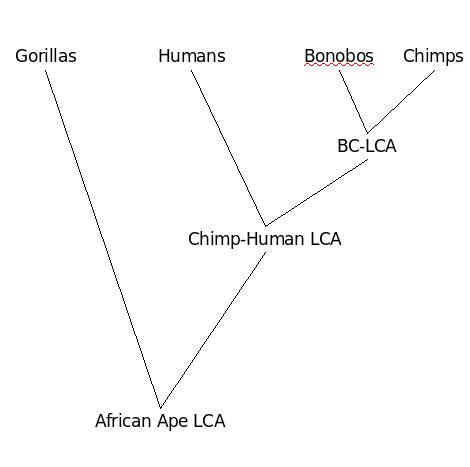
نسل غير منسجم
وثمة
جانب آخر مثير للاهتمام هو أنه ERV لا تظهر دائما الانماط المتوقعة
التطورية "الميراث". وفقا لشجرة النشوء والتطور المقترحة الشمبانزي هي
أقرب إلى البشر من أن الغوريلا. ونظرا لهذا السيناريو، فان الغوريلا
والشمبانزي فقط من المتوقع ان تتشارك ERV الا إذا كانت هذه ERV نفسها كانت
موجودة في البشر. ولكن هناك بعض فيروسات النسخ العكسي التي يبدو انها لا
تظهر هذا النمط. على سبيل المثال، عائلة K من ال ERVs
(HERV-K provirus) موجود في الشمبانزي والغوريلا، ولكن ليس في الانسان، أجزاء من ERVs المعروفة باسم CERV 2 و CERV 1
هي
عناصر موجودة في الشمبانزي، الغوريلا والبونوبو (غير متقابلة ) لكنها
غائبة في الإنسان، انسان الغاب، قرود العالم القديم، قرود العالم الجديد .
إن التفسير المعتاد لهذه النتائج، بطبيعة الحال، هو أن البشر فقدوا هذا أو
ذاك ERV بصورة خاصة على طول الطريق. بالطبع، يمكن أن تستخدم هذه الحجة
يمكن ان تستخدم لتفسير اي اختفاء. يبدو من الصعب أن نتخيل إلى حد ما، ومع
ذلك، كيف يمكن لمجموع التجمع السكاني البشري أن تفقد ERVs التي يتم
الاحتفاظ في كل من الشمبانزي والغوريلا؟ - بعيدا عن فكرة عنق الزجاجة
للتجمع السكاني .
هناك أيضا مشاكل اكبر في التسلسل النسلي غير المنسجم لفيروس النسخ العكسي :
"أجرينا تحليلين لتحديد ما إذا كانت هذه ال12 فترة خريطة مشتركة
هي في الواقع انفصلت بعد تكوين الانواع. أولا، فحصنا توزيع المواقع
المشتركة بين الأنواع (الجدول S3). لقد وجدنا أن التوزيع يتعارض مع النسالة
المقبولة عموما في الرئيسيات سفلية المنخرين. هذذا مهم على وجه الخصوص
بالنسبة لخط الانسان / القردة الكبيرة .على سبيل المثال، تتم مشاركة فترة
واحدة فقط من قبل الغوريلا والشمبانزي، في حين، يتم مشاركة فترتين من قبل
الغوريلا والبابون، في حين تتشارك وبصورة ماثلة للعيان المكاك والشمبانزي
في ثلاث فترات .تحليلنا الجنوبي يدل على أن الإنسان وانسان الغاب تفتقر
تماما لتسلسل PTERV1 (انظر الشكل 2A). إذا كانت هذه المواقع انفصلت بعد
تحديدي االانواع حقا، وبالتالي يوجد لها اسلاف في جد البشر / القرد، فانه
يتطلب ان يتم حذف ستة مواقع في خط نسب الانسان . وعلاوة على ذلك، فإن
بالضبط نفس الستة مواقع كانت لتحذف في انسان الغاب وبنفس الطريقة لو ان
تسلسل النسب المقبول بصورة عامة كان صحيحا . مثل هذا المسلسل من أحداث
الحذف المستقلة في مواقع محددة من الجينوم من غير المرجح حصولها (الشكل
S3).
خطوط عديدة من الأدلة تشير إلى أن نسخ PTERV1 في الشمبانزي
والغوريلا نشأت من مصدر خارجي . أولا، ليس هناك عمليا أي تداخل (أقل من 4٪)
بين مواقع الإدراج في كل من الشمبانزي، الغوريلا، المكاك، والبابون، مما
يجعل من غير المرجح أن نسخ الفيروس الداخلي موجودة في سلف مشترك ومن ثم حدث
لاحقا حذف في خط نسب الإنسان وانسان الغاب . ثانيا، شجرة النشوء والتطور
للPTERV1 لا تتفق مع شجرة الأنواع المقبولة عموما للرئيسيات، مما يشير إلى
انتقال الأفقي بدلا من الانتقال العمودي من سلف مشترك . قد يكون هناك تفسير
بديل أن شجرة نسل الرئيسيات غير صحيحة إلى حد بعيد، كما اقترح من قبل
أقلية من علماء الأنثروبولوجيا ".
"التناقض مع تحليلات النشوء والتطور موجود فعلا وغالبا ما يفسر في كثير من الأحيان بالحجج الدبقة بدون دليل ايجابية".
في
الواقع، يبدو انه يمكن تفسير أي حقائق أو مجموعة بيانات داخل النموذج
التطوري باستخدام الحجج هذا أو ذاك "المخصص" لجعل البيانات تناسب النظرية.
هذا ينتج مشكلة التحيز عندما يتعلق الأمر بتفسير مجموعات البيانات. تمت
ملاحظة البعض من هذه التحيزات في شجرة نسل فيروس النسخ العكسي. على سبيل
المثال، وفقا لبوسادا وكراندال، في ورقة 2001 المنشور في البيولوجيا
الجزيئية والتطور:
"نماذج خاطئة من تطور [الفيروسات العكسية] قادت
الى تقدير الأشجار التي هي في اتفاق مع الأدلة البيوكيميائية والمناعية
ودراسات النشوء والتطور مع السابقة ...
عند النظر في نتائج هذه
الدراسة، فقط تلك الأشجار والتي قدرت وفقا لنماذج التطور البسيطة والمحتمل
ان تكون خاطئة فقط تتفق مع الأدلة الحالية. في معظم لأشجار اللتي أعيد
بناؤها، ألاجناس المختلفة تظهر كمجموعات متفردة. هذه المجموعات لديها قيم
عالية التمهيد عادة تشير إلى أنه، بالنظر إلى مجموعات البيانات اللتي في
متناول اليد، نحن يمكن أن يكون على ثقة في العقد اللتي تحدد هذه المجموعات.
عندما يتم توظيف نماذج تطور أكثر تعقيدا، وأكثر واقعية، يتم استرداد
مجموعات أجناس اقل كمجموعات متفردة ، ويصبح مستوى الدعم أقل، والطبولوجيا
مختلفة جدا من الاشجار " المعروفة "المفترضة.
الانحياز في الانسال
والذي من خلاله تعطي النماذج الخاطئة اجابات صحيحة تم التعرف عليه في
دراسات محاكية . لماذا يحدث هذا الانحياز يبقى هذا السؤال غير محلول .
عامل
محتمل ممكن ان يساهم في هذا الانحياز هو التراصف المعضل والذي يحدث هو انه
تسلسلات مجموعة (جنس ) معينة تتراصف بكل سهولة بينما يحصل العكس للمجموعات
المختلفة . النماذج المعقدة ممكن ان تكون مرتبكة عندما نحاول استخراج
المعلومات من تراصف تسلسل داخل المجموعة السيء . هذا من شأنه أن يضمن كلمة
تحذير لتقدير الانسال من مجموعات البيانات المتباينة للغاية.

العدد الكلي لفيروسات النسخ العكسي
منذ
وقت ليس ببعيد كان يعتقد أن حوالي 30،000 فيروس نسخ عكسي موجودة داخل
الجينوم البشري / القرد، تمثل حوالي بين 1-8٪ لكل منهما. كما ورد في اجتماع
سلسلة وتحليل جينوم الشيمبانزي لعام 2005 ، حيث تمت مقارنة جينوم
الشيمبانزي بأكمله إلى الجينوم البشري، ويعتقد الآن أن ما يقرب من 200،000
ERV، أو أجزاء من ERV، توجد داخل الجينوم في كل من البشر والقردة - بلغ
مجموعها حوالي 127 مليون زوج قاعدي (حوالي 4٪ من مجموع التركة الحقيقية
الجينية) بعض الكتاب يشير إلى وجود أصل ERV 45٪ للجينوم البشري بأسره
(ميندل وماير 2001) و 50٪ لأنواع الثدييات بشكل عام ، إذا ما تم تضمين كافة
ألاجزاء الصغيرة من تسلسل ERV في التقدير. وفي اي حالة من هذه المئات من
الآلاف من الأجزاء المعروفة من ERV، فإن الغالبية العظمى منهم يبدو ان
تتقابل، في المواضع نفسها، بين البشر والشمبانزي. كما اقترح في "جدول 2" من
الورقة ، وأقل من 1٪ من ERV هي خاصة بخط النسب الخاص سواء للإنسان أو
القردة. وبعبارة أخرى، يتم مشاركة الغالبية العظمى من ERV أو "انها انفصلت
بعد تكوين الانواع " بين البشر والشمبانزي (زيادة كبيرة من سبعة أو قبيل
ذلك بحيث كان يعتقد مرة واحدة لتصيب كل من البشر والشمبانزي في مواقع
مماثلة ،ملخص ERVبالنسبة للعديد فان هذه النتيجة وحدها(من التماثل العالمي
تقريبا ل ERVs بين الأنواع) قد تشير الى أدلة دامغة لصالح أصل مشترك -
ناهيك عن أنماط التداخل الهرمية بين الأنواع التي تظهرها معظم هذه
الفيروسات . بالتأكيد، لا يبدو منطقيا إنكار ألاصل المشترك نوعا ما. ومع
ذلك، وبالنظر إلى الاكتشافات المتزايدة لوظائفية العناصر المختلفة لERVs،
في أشكالها المختلفة وشظاياها، جنبا إلى جنب مع صعوبة شرح كون وجودها من
مصادر خارجية حقا فان نظرية أصل تطوري مشترك يصبح أقل يمكن الدفاع عنه.
وجود ERVsأو أجزاء منها في نفس الأماكن أو الاماكن الشبيهة في الغالب
واللتي تقوم غالبا بنفس الاعمال خلال الجزء الاكبر من الجينوم (غالبا ما
تكون على مستوى عال جدا من التعقيد المتكامل) في المخلوقات المماثلة تؤيد
بقوة نظرية التصميم المشترك.
الآلية الداروينية من الطفرات العشوائية
والانتقاء الطبيعي المستند الى الوظيفة هي ببساطة غير كافية لشرح التعقيد
الوظيفي رفيع المستوى للعناصر داخل جينوم الERV المعقدة جدا والمتكاملة
بشكل كبير. و أيضا، الحقيقة القائلة بأنه ببساطة لا توجد أي أمثلة معروفة
على فيروسات نسخ عكسي خارجية قامت بإدراج نفسها في الخطوط الجرثومية لكل من
البشر أو الشمبانزي، أو أي حيوان آخر في هذه المسألة. إذا كانت فرضية
الأصل المشترك هي في الواقع صحيحة، فأن مثل هذه الأحداث وقعت في الماضي
بمعدلات سريعة للغاية، وينبغي للمرء أن يتوقع له بأن يرى ما لا يقل عن بضعة
أمثلة من هذا القبيل في الوقت الحالي . فلماذا لا توجد هذه الأمثلة
بالفعل ؟ الشروط الاساسية لحصول عشرات ومئات الآلاف من الإصابات الفيروسة
المحايدة لتحقيق التثبيت في ألخط الجرثومي الأصلي في كل من البشر
والشمبانزي، مما يتطلب اختناقات سكانية صغيرة جدا على مدى فترات طويلة من
الزمن، هو أيضا غير متناسقة بشدة مع فرضية الانحدار المشترك على العديد من
المستويات، في حين يجري بصورة صحيحة وفقا لفرضية التصميم الذكي الأصلية مع
التغيرات الانحلالية تعمل على مدى فترات لاحقة من الزمن.
ولكن لماذا أي
مصمم ذكي ان يقوم عمدا بتصميم عناصر فيروسات كجزء من الجينوم للكائنات
الحية شديدة التعقيد؟ اليس إدراج الفيروسات العكسية ضار وظيفيا دائما
تقريبا؟ بالنظر إلى أنه من المعروف الآن أن الكثير من الميزات المفيدة
مرتبطة بهذه عناصر الفيروسات الداخلية ، جزء كبير من جينوم الانسان في
الواقع تحت سيطرة هذه العناصر، وأننا لن نكون قادرين على العيش بدونها،
يبدو من المحتمل جدا أن الجينوم الأصلي كان قد تم تصميمه مع هذه العناصر في
مكانها المعروف منذ البداية المبكرة -بحيث انها كانت دائما حيوية لوجود
البشر والقرود، وجميع الكائنات الحية المعقدة الأخرى. ومع ذلك، كما هو
الحال دائما عندما تتعرض الالات التكاثرية المعقدة أو أسطر التعليمات
البرمجية للطفرات العشوائية والانتقاء الطبيعي،التغيرات التنكسية تحدث
حيثما يتم ادراك العناصر الطفيلية بصورة سريعة من خلال فقدان ما موجود
مسبقا من التعقيدات التي تستخدم للسيطرة على أو تعديل أنشطتها . بل ما هو
أبعد من ذلك ، أن فيروسات النسخ العكسي الخارجية التي يعاني منها الانسان
حاليا (مثل فيروس نقص المناعة البشرية، HTLV-1، التهاب الكبد B-، الخ)
المستمدة في الأصل من تسلسلات فيروسات نسخ عكسي داخلية و التي عانت تغيرات
الاضمحلالية. مثل السرطان القلة التي نجت من نظم الرقابة والتنظيم
الاعتيادية ، أصبحت هذه الفيروسات الخارجية "أنانية" وطفيلية، وتهاجم
وتتغذى على مضيفها بدلا من المساهمة في وظيفة مثالية للمضيف.
هناك
العديد من الأمثلة في الوقت الحالي على مثل هذه التغيرات الاضمحلالية التي
تؤدي إلى وظائفية طفيلية - مثل نظام حاقن السم TTSS في البكتيريا التي
تطورت من خلال فقدان عناصر تركيبة موجودة سابقا في السوط الدوار البكتيري .
ويستخدم الآن نظام TTSS من قبل البكتيريا السامة - مثل البكتيريا التي
تسبب الطاعون الدبلي أو "الموت الأسود" (Yersinia pestis). والتي من الافضل
ان يشار إلىها على انها تغيرات عكس تطورية على انها تطورية لانها تتضمن
فقدان لوظيفة وليس اكتساب لتعقيد وظيفي جديد.
كرمسوم 2 كنا قد بينا في موضوع سابق مدي تخبط الدروانية
في هذا الموضوع
نكمل باقي البحث في موضوع كرمسوم 2
إنه قول شائع أن النسخة البشريّة من الكرموسوم 2 عانت - حقيقةً -
من حدث إندماجى فى وقت ما بالماضى (أو سابقا ) ؛فالتيلوميرات كمثال ، عادةً
ما تُوجد فقط فى نهاية الكروموسومات ، وليس فى المُنتصف ، ولكن فى مُنتصف
الكرموسوم 2 هناك بالترتيب سلسلة مُتعاقبة ( أو تسلسل من البريتلوميرز
(pretelomeric )، فتسلسل من التيلوميرز (telomeric)
، فتسلسل من التيلوميرز مقلوب(inverted telomeric )، فتسلسل مقلوب من البريتلوميز(pretelomeric ) ،
هناك
أيضا بقايا من السنترومير الزائد (أو الإضافى ) ، كما أن هناك أنماط للربط
مُتشابهة مع الكروموسومات المُكافئة (2p and 2q)في القردة العُليا
لذا فإنّه يبدوا واضحا تماما لغالبية أنه كان هناك حدث إندماجى كروموسومى فى وقت ما بالماضى ، فى سلف الإنسان الحديث ،
ولكن
حَدَثُ الإندماج - الذى نتج عن تكوين الكروموسوم البشرى الحديث 2 - ليس
ليس أبدا فى تعارض مع نظريّة السلف المُستقل ( المُنفصل ) للإنسان والقردة (
أو القردة العليا ) ، وذلك على الرغم من التأكيد الشائع جدا من قبل التيار
السائد من العلماء، والعلماء المُدافعين عن الداروينيّة أن إندماج
الكرموسوم 2 دليل واضح جدا على السلف المُشترك بين الإنسان والقردة ( أو
القردة العُليا ) .
كيف امكنني حتى ولو فرضا ان اقترح هكذا حجة بالضد من التيار السائد للعلماء ؟
حسنا ، لشئ واحد : الإندماج الكروموسومى هو امر شائع إلى حد كبير - حتى فى نفس النوع .
فى
الحقيقة ، هناك بشرُ أحياء اليوم ، عندهم إندماج كروموسوى ،( أى بل )
وتعجب وتعجب ، إنّهم ما زالوا بشر ! لايُمكن تمييزهم عن غيرهم من البشر
المُعاصرين شكليا ووظيفيّا .
مثال آخر يُمكن أن نجده فى الخيول ، الخيل
البريّة الهجينة لها 33 زوج (أى من الكروموسومات ) بينما الحصان المُستأنس
( أو الأليف ) به 32 زوج من الكروموسومات ،
أيضا الكلاب المنزليّة ( أى
المُستأنسة ) والذئاب التى من جنس الكلب بها 78 كروموسوم بينما الثعالب
بها عدد من الكروموسومات يتراوح بين الـ 38 - 78 .
بل ومثال آخر ، هو
الفأر المنزلى ( Mus Musculis ) الذى به 40 كروموسوم ، بينما غالبيّة
الفئران التى شكلت فئران جبال الألب الإيطاليّة ( أى هى عند جيال الألب )
وُجد أن بها فقط 22 كروموسوم. ( رابط : http://www.detectingdesign.com/donkeyshorsesmules.html
لذا ، فإنّ إختلاف عدد الكروموسومات بين الإنسان والقردة ( أو القردة العُليا ) لا يُشير بالضرورة إلى سلف مُشترك ،
إنه
ليس دليلا على متى وقع هذا الحدث ، ولا دليلا على سلف سابق لهذا الحدث
.انّه ببساطة يعنى أن المخلوقات المُتشابهة ، مع ( كونها لها ) سلف مُستقل
من الأصل ، هذه المخلوقات لها نفس عدد الكروموسومات ، و( نفس ) النمط
العام للربط ( بين الكروموسومات ) ، ( هذا ) العدد الذى تغير لاحقا
بالطفرات الإندماجيّة فى البشر أثناء الإختناق السُكّانى أوعندما بلغ عدد
السكان عنق الزجاجة تكدسا ) وبإعطاء احتمال حصول أختناق سُكانى مُثير آخر
فى المُستقبل ، (فإنّ ) مثل هذا الإندماج المُتنقل يمكن أن يحدث بسهولة مرة
ثانية ، فى أى من القرود (أو القردة العليا ) أو الأناسي .... أو أى مخلوق
آخر بالنسبة لهذه المسألة .
هذا ما يُمكن أن يُتوقع بوضوح هنا ، حتى
أوُلئك الذين يعتقدون فى التصميم الذكى(ID) يفهمون أنّه ليس كل السمات
الوراثيّة تتطلب مُساهمة الذكاء .الإندماج البسيط للكرموسوم الثانى - بدون
إكتساب أو فقد أى وظيفة مُهمة - من السهل تفسيره عن طريق العمليّات
العشوائيّة الطائشة ( أو الغير عقلانيّة ) و هى فى الواقع مُنتشرة إلى حد
كبير .
ليس بالامر المهم جدا . و ليس شيئا مُفاجئا جدا أو صادما -
ولا حتى من وجهة نظر القائلين بـ ID .فى الواقع ، التطوريون سيستمروا
بالجدال حول السلف المُشترك للإنسان والقردة (أو القردة العليا ) دون
الأخذ ـبإندماج الكرموسوم 2 .
هذا الحدث الإندماجى يُضيف - حقيقتا - لا شئ للجُحّة .
إنّه
ببساطة لا يُقدّم تفسيرا إضافيا أو قوة تنبُئيّة للمُنازعة حول السلف
المُشترك ، شيئا سوى الملاحظة البسيطة حول وجود سلف من نوع ما بناءا على
التشابه بين الانسان والقرد
بعبارة أُخرى ، هؤلاء التطوريون الذين عرضوا
هذه الحُجة لا يُقدّمون أى دليل على أن سلف الإنسان - الذى كان لديه فى
الأصل 48 كروموسوم( كما موجود عند القردة) - له " علاقة " حقيقيّة وثيقة
بالقردة تجعله يبدو شكلا ووظيفتا شبيه بالقرد . .
إنّها ليست حقيقة كونّ القردة بها 48 كروموسوم ، تجعلها تبدو وتتصرف و تؤدى مثل القردة وليس كالبشر .
لو
كانت المسالة بهذه البساطة ، لكان التطوريون يملكون - حقيقة - حجة قويّة
جدا .المُشكلة عند الداروينيين هى ان الموضوع ليس بهذه البساطة ولاحتى قريب
منها ) .إختلاف عدد الكروموسومات هذا ، فى حد ذاته ، لا ينتج عنه أى
إختلافات وظيفيّة واضحة بين القردة والإنسان _ على الإطلاق
ولذا ،
فإنّه ليس تمددا أن نفترض بأن أى سلف للإنسان الحديث ذو 48 كروموسوم ربما
كان له إيضا مُخطط كروموسومى شبيه للقردة ، بغض النظر عن إذا ما كان هذا
الفرد له " علاقة " بالقردة عن طريق سلف مُشترك أم لا .
فالإدعاء بأن
نمط الربط المُتشابه دليل على سلف مُشترك مع القردة ، ببساطة يستدعي الفكرة
القائلة بأن " التشابه = سلف مُشترك " ، وبالتالى يطرح السؤال .
فى
الوقت الذي يبدو فيه من المعقول تماما أن أوجه التشابه القويّة جدا ، مثل
تلك الموجودة بين الإنسن والقردة العليا ، تُشير واقعيّا إلى سلف مشترك ،
فإنّ هذا السلف المُشترك ليس بالضرورة مُستندا إلى تحدر مُشترك من بعض
الأسلاف المُشتركة عن طريق تعديلات جينيّة بطيئة أُختيرت على مدار الزمان
بواسطة طبيعة طائشة ( غير عقلانيّة )
.
فإنّه نظراً للإختلافات
الوظيفيّة عالية التعقيد بين النوعين ( الإنسان والقردة ) ، والتى أُكتُشفت
أكثر وأكثر فى السنوات الأخيرة (( خصوصا فى المناطق الغير مُرمّزة
non-codingمن الجينوم )) ؛ فإنّه يبدوا أكثر أحتمالا أن السلف المُشترك
لهذه الإختلافات ، وكذلك أوجه التشابه ، مُستندا إلى تصميم مقصود غاية فى
الذكاء .
الحدث (( الأحداث )) الوحيدة التى لا تحتاج - بوضوح - إلى
تدخل ذكاء خارجى ذو مستوى عالى ، هى الأحداث مثل الإندامج الكروموسومى
العشوائى أو غير ذلك من أشكال الطفرات العشوائيّة ، والتى هى بعيدة
الإحتمال جدا أن تُنتج ( هذه الطفرات العشوائيّة ) أىّ إستفادة وظيفيّة
تتجاوز المستويات المُنخفضة جدا من تعقيد الوظائف .
( رابط :
مرة
أُخرى ، إنّه من الممكن جدا ، بل من المُحتمل جدا حقيقة ، أنّ البشر
أسلافنا خضعوا لحدث إندماجى كروموسومى أثناء الإختناق السُكّانىّ ، فى
التاريخ الحديث الى حدّ ما (( أى فى غضون عدة آلاف ، من السنوات الماضية،
على الأكثر )) ، ( وهذا ) يُفسّر بسهولة إندماج الكروموسوم2 .
ويُدّعم هذا المفهوم مقالُ نُشر فى العدد 2003 من ( مجلة ) الطبيعة ،
بواسطة رود وآخرين عندما طرح الكاتب الحُجة التاليّة :
تُشير
هذه التحليلات إلى أنّ سلاسل نسب جميع البشر الأحياء تداخلت (أو تراكبت )
بطريقة ملحوظة فى الماضى القريب .وعلى وجه الخصوص الـ MRCA ( أحدث سلف
مُشترك ) لكُل البشر فى الوقت الحاضر الذي عاش منذ بضعة آلاف سنة مضت فقط
(( تقريبا 3000 سنة)) فى هذه النماذج .
علاوة على ذلك ، ( فإنّ ) جميع
الأفراد الذين عاشوا قبل الـ إم آر سى آيه ببضعة آلاف سنة فقط ،فان كل
إنسان فى الوقت الحاضر لديه بالضبطا نفس مجموعة سلاسل أنساب الأسلاف .
التسلسل الخلالى التيلوميرى
وفى هذا الخط أيضا ، فإنّه من المُثير للإهتمام أن نُلاحظ أنّ التسلسل التيلوميرى " الخلالى " ITCS
، مع تسلسله المتكرر TTAGGGTTAGGGTTAGGG ( الموجود ) بهم
.... وُجدوا مُبعثرين ( أو مُتناثرين ) فى جميع أنحاء الجينوم البشرى
والقردة العليا _ فى كل مكان . كان دائما يُعتقد أنّ هذا التسلسل الخلالى
هو ببساطة تسلسل خُردة ، خلّفتها القمامة التطوريّة فى العصور الماضيّة .
ولكن
، إنّه _ منذ ذلك الحين _ أُكتشف أن هذه الـ ITSsغالبا ( ما تكون ) مُهمة
وظيفيّا للجينوم .فالتنظيم الصبغي (الكروماتيني ) للتيلميرات يُمُكنه أن
يُسكت الجين ( عن العمل ) وقد تم ربطه بنمط الوراثة فوق الجيني .
وعلاوة
على ذلك ، فإنّ مُختلف فئات النسخ مُشتقة من التيلوميرات ، و من مناطق الـ
دى إنّ إيه المُرافقة ( أو المُحيطة ) المُتكررة الخاصة بهم . ( .
وأيضا يُشاركون فى العديد من الوظائف الخلويّة والتنموية (للكائن )
ما هو مُثير للإهتمام أيضا هو أنّ هناك العديد من تسلسلات الـ
ITS المعروفة فى جينوم البشر والقردة العُليا واحد منهم فقط، عند مكان
2q13، هو المُشترك بين الإنسان والشمبانزى .وبعبارة أُخرى ؛ فإنّه من الـ
ITS
العديدة والمعروفة فى جينوم كُلّ من البشر والقردة العليا (( والبقر ،
والدجاج والفئران .. إلخ )) ،( من كل هؤلاء ) يُمكن فقط للـ 2q13 أن يكون
لها إرتباط بنقُطة تطورية أو حَدَث الإندماج بينما الـ ITS الأُخرى لا تصطف
كنقاط تطورية كروموسوميّة فى الرئيسيات .لذا ، فإنّ القول بأنّ2q13 ITS
هو نموذج مُطابق لما يُرى فى خلال كُلّ من جينومي البشر والشمبانزى ، يبدوا وكأنّه انتقاء للحقائق المُتاحة .
معظم تسلسلات الـ ITS المعروفة ليست " ندبات الحمض النووي " بالطريقة التى صُوّرت بها بل على الأصح ، فإنّه يبدو أكثر احتمالا أنّ
ITS هى الأماكن التى أُضيفت فيها مُكررات TTAGGG ببساطة للكروموسومات بـواسطة إنزيمات التيلوميريز ، وأنّ عديد من أماكن الـ
ITSمُرتبطة بمجموعات مُتميزة من البروتينات والتى هى ( بدورها ) مُرتبطة
بأدور وظيفيّة هامة داخل الجينوم _ مثل إعادة تركيب النُقط الساخنة (أو
المناطق الفعّالة ) ، إلخ .
نموذج ( أو مثال ) جديد
نمط الحياة الهندسى المتكرر و المُعقد
فى
ضوء المعلومات المُقدّمة حتى الآن ، فإنّه يبدوا واضحا تماما أنّ الفكرة
القديمة ، والتى تقول أنّ الجينات الكاذبة وغيره من أشكال الـ دى إنّ إيه
المُشتركة " الخردة " تُعطى دليلا واضحا على أصل مُشترك ، ( والتى هى )
أكثر شيوعا من ( أنّ لها ) ضرورة وظيفيّة ،- هذه الفكرة - لابد من التخلص
منها بجانب ( التخلص من ) فكرة أنّ مُعظم الجينوم البشرى هو عبارة عن
قمامة تطوريّة ، لا وظيفة لها .
بالتأكيد ، لو تشاركت الكائنات نفس
البيئة ، وكان عندهم تشابه فى المظاهر الشكليّة والإحتياجات ، فليس للمرء
أن يتعجب جدا عندما يجد عناصر وظيفيّة جينيّة مُماثلة ، مُشتركة بين هذه
المخلوقات .
لذلك ، هذه التسلسلات لا يمكن أن تُستخدم لتؤسس بوضوح
الشجرة التطوريّة أو لتقدير الأوقات ( أو المرّات ) المُختلفة أى منذ تم
المُحافظة على مثل هذا اللتسلسل المُفيد عن طريق الإنتخاب الطبيعى دون أى
تغيرات مُهمّة . .أوجه التشابه والإختلاف لن تستند كثيرا على التغيرات
التطوريّة على مدار الزمن نظرا لمُشاركة سلف مُشترك ، لأنهم ( السلف
المُشترك ) سيكونوا ( عندئذ ) نتيجة لأوجه التشابه والإختلاف فى الإحتياجات
الوظيفيّة ( ولكنّها ) التى كانت دائما موجودة هناك ، و مُحافظ عليها من
قوة الإنتقاء الطبيعى ، ( وذلك ) مُنذ أن أتت تلك المخلوقات .
ولكن
وراء هذه الحُجة الأساسيّة (شئ مُهم )هو الإكتشاف الرائع لحقيقة مُعظم الـ
دى إنّ إيه الغير مُشّفر فى الجينوم البشرى ، ( و أنّه ) ليس خُردة (
ناتجة ) بعد كُلّ شئ .
فى الواقع ، هو بلا جدال الجزء الأكثر وظيفة فى الجينوم ، يُوجّه موضع الطوب والأساسى والملاط للكائن الحىّ _ إذا جاز التعبير .
ويُجرى إكتشاف طبقة من المعلومات فوق طبقة من المعلومات .
كما
هو الحال فى النمط الهندسى المُتكرر ، ( فإنّ) كلما أفترب نظر المرء ( منه
) ، كلما ظهرت تفاصيل وتعقيد وتركيب العناصر الوظيفيّة للجينوم بوضوح أكثر
، طبقة بعد طبقة
NCODE (ENCyclopaedia of DNA Elements)
لا
أحد حتى الآن يعلم ولو حتى مُجرّد ما ستبدو عليه الصورة الكُبرى للجينات
(أو لعلم الوراثة ) إذا أصبحت هذه الطبقة الخفيّة من المعلومات مرئيّة .
"
فى الواقع ، ما كان ملعونا وخُردة ( أو غير مرغوب فيه ) لكونه لم يُفهم ،
ربما حقيقة يتحول إلى أن يكون أساسى للغاية فى ( معرفة ) التعقيد الإنسانى "
.أشار إلى ذلك ماتتيك .
( فالبسبنة للجينات الزائفة ، الريبوسويتشس ( أو رموز تبديل الريبو ) ، وكلّ ما تبقى جانبا فان هناك سبب جيد للشك فى أنّ هذا صحيح .
ال
آر إنّ إيه النشط ، إنّه الآن بدأ يُعرف ، فهو يُساعد فى التحكم فى
الأبنية واسعة النطاق فى الكروموسومات ، وبعض التعديلات الكيميائية الحاسمة
لهم _ والتى تُغير كُليّا طبقة المعلومات اللاجينيّة ( أو الجينيّة ) فى
الجينوم . 16
فى الحقيقة ، الفهم الأكثر تفصيلا _ حتى الآن _ فى
كيفية عمل الجينوم البشرى قادت العلماء إلى أن إعتقاد ( أو إبرام ) (( كما
فى 14 يونيو 2007 )) أن مفهوم حجر الزاوية حول الرموز الكيميائية للحياة
هو إعتقاد معيب ( أو خاطئ ) بشدة .
التقارير فى مجلة الطبيعة
البريطانيّة والجريدة الأمريكيّة لأبحاث الجينوم يوم الخميس (( 14 يونيو
2007 )) ، هم يُشيرون إلى أن أىّ نظرية أُنشئت عن الجينوم ينبغى أن تودع
فى التاريخ بين الجينات والتسلسلات المعروفة بتنظيم نشاطهم ، إمتدادات
طويلة ( و) مُملّة والتى يبدوا أنها لا تفعل شيئا
المُصطلح المُستخدم
لهم هو دى إنّ إيه " خُردة " ، مما يعكس إفتراض أنّهم مُجرّد أخشاب طافية (
لا قيمة لها ) من ماضينا التطورى ، و ( أنّهم ) ليس لهم وظيفية بيولوجيّة
.، ولكن العمل ( المبذول ) بواسطة تحالف (أو إتحاد ) الـ NCODE يعنى أنّ
مفهوم الشذرات من التفاهة _ هذا _ عن الـ دى إن إيه ينبغى أيضا أن يُلغى
الجينوم تبين أنّه آلة مُتشابكة ( أو مُتداخلة ) ، مُعقدة للغاية ، مع وجود إمتدادات غير نشطة قليلة جدا ، أقرّه الباحثون
الجينات ، تُبيّن ( أو تستشف أى منها ) ، هى واحدة فقط من الأنواع العديدة من تسلسلات الـ دى إنّ إيه ، لها دور وظيفيّ .
و
الـ دى إنّ إيه " الخردة " تبيّن أنّ له دور أساسى فى تنظيم عمليات صناعة
البروتين .,واللتي سابقا شُطبت كصامتة ( أى لا وظيفة لها مُكتشفة ) ، الآن
خرجت كمغنيّة لها صوتها الخاص السرى (أو الحكيم ) و كـجُزء من جوقة
التفاعلات الجُزيئيّة الواسعة .
يتم نسخ الغالبيّة العُظمى من الجينوم
أو كتابتهم فى آر إن إيه ، الذى هو الجُزى الفعّال فى خلايانا ، ( فهو )
يُبادل (أو يُرحّل ) المعلومات من دى إنّ إيه المحفوظة ( أو الارشيفيّة )
إلى الآلات ( أو المكينات ) الخلويّة ( أى التى بالخلايا ) "
قاله تيم هوبارد من معهد ويلكوم ترست نسانجر ، ( الذى هو ) مجموعة أبحاث بريطانيّة كانت جزءا من الفريق .
" هذا إكتشاف رائع ، حيث أنّ أكثر البحوث السابقة تفترض أن جزء من الجينوم فقط ( هو ما ) يمت كتابته "
وقال
فرانسيس كولينز ، مُدير العهد القومى الأمريكى لبحوث الجينوم
البشرى(NHGRI), ، والذى حشد 35 جماعة علميّة من جميع أنحاء العالم فى
مشروع الترميز ، قال : المُجتمع العلمى " سيحتاج أن يُعيد التفكير حول
بعض وجهات النظر ذات الأمد الطويل والتى كانت حول ماهيّة الجينات و ما
تفعل " 17
" نحن خدعنا انفسنا بالتفكير فى أنّ الجينوم كان على
وشك أن يكون خطة شفافة ( أو مُخطط شفاف أى يتسم بالوضوح ) يقوله مل جريفز ،
وهو عالم أحيائى فى معهد أبحاث السرطان فى سوتون ، بالمملكة المتحدة .
بدلا
من ذلك _ كما فى التسلسل والوغيرها من التكنولوجيّات الجديدة تبث البيانات
عليها _ ( فإنّ ) تعقيد البيولوجيا يبدوا ناميا من حيث الحجم ( أو وفقا
لأهميتها ( أى أن العلوم تُبسط وعلم البيولوجى يزداد تعقيدا )
وكان
الخوض فيه مثل تكبير ( الزوم على ) مجموعة ماندلبروت _ التى هى مساحة يتم
تحديدها بمعادلة بسيطة _ ولكن هذا يكشف عن أنماط أكثر تعقيدا من أى وقت
مضى مثل أو لأنّ واحد من الأقران قريب إلى حدودها
" إنّها تبدوا
وكأننا نتسلق جبلا ، ( بينما هو ) يظل ( إرتفاعه ) أعلى فأعلى " تقوله
جينفر دودنا ، عالمة بالعكسماء الحيويّة فى جامعة كاليفيورنيا ، بيركلى .
" كلما إزدّنا معرفة ، كلما أدركنا أن ( هناك ) ما يُمكن معرفته "
أظهر
باحثون من مشروع تعاونى دولى ن يُسمّى موسوعة عناصر الحمض النووى ((إنكود
)) أنّه فى جُزء محدد من الجينوم ( ظهر أنّه ) يحتوى على نسبة مئويّة قليلة
فقط من تسلسلات ترميو البروتين ، بين 74 % ، 93 % من الـ دى إنّ إيه تمّ
كتابتها ( أو نسخها ) إلى آر إنّ إيه
مُعظم الـ دى إنّ إيه الغير مُرّمز
له دور تنظيمى ، الـ آر إنّ إيه الصغيرة المختلفة الأنواع يبدوا ( أنها )
للتحكم فى التعبير الجينى على مستوى نسخ (او كتابة ) كلُ من الـ دى إنّ إيه
و آر إن إيه بطرق ، والتى ( الطرق ) ما زالت فقط فى بدايتها لتكون واضحة .
فقط
الوجود المطلق ( أو الكامل ) لهذه المُتظِمات الغريبة تُشير إلى أنّ فهمنا
عن الأشياء الأساسيّة _ مثل كيف يمكن تشغيل وإيقاف الخليّة _ ساذج بشكل لا
تُصدق " يقوله جوشوا بلوتكين ، عالم الأحياء الرياضيّة فى جامعة بنسلفانيا
بفينلندا . 57
الجينوم البشرى بالأرقام . 26
* 1.5 % من الجينوم تُرجمت إلى بروتين .
* 27 % من الجينوم كُتب ( او نُسخ ) كجزء من التعبير الجينى لترميز البروتين ، ولكن لم يُترجم إلى بروتين .
* 25 % من الجينوم الذى تمت كتابته كتابته ( او نسخه ) ولكم لم يُترجم ، وليس له إرتباط بجينات ترميز الربوتين .
* 250 ميكرو آر إنّ إيه حُددت حاليا ( فى يونيو 2005 ) .
، تقريبا 1.000 ( كما فى 2007 ) ( رابط :
http://www.biomedcentral.com/1472-6750/7/36
*
10.000 جينات ترميز البروتين أُعتبرت مُنظمة بواسطة ميكرو آر إنّ إيه ،
كل ميكرو آر إنّ إيه يمكنه إستهداف عدة جينات ، كما أنّ جين مُعين ربما
يُتظَم بواسطة عدة ميكرو آر إنّ إيه .
* 98 % من الناتج الجينى هى آر إنّ إيه غير مُرمّز .
* 9 % من الجينات التى يظهر أنّها يصحبها جزيئات مضادة للانتساخ ( اى نسخ ) النصوص .
* تقريبا 20.000 " جينات كاذبة " فى الجينوم .
هذامُثير للإهتمام جدا ، أعنى ، من كان يظن انّ أغلبية الجينوم نُسخت او كُتبت إلى آر إنّ إيه ؟ _ وسيكون لها وظيفة حقيقة ؟
فقط
منذ بضع سنوات مضت ، المُجتمع العلمى إعتقد بأنّ أقل من 5 % من الجينوم له
وظيفة واقعيّا ، و أنّ البقيّة هم بقايا تطوريّة غير وظيفيّة ( أى لا
وظيفة لها ) .
بعد كل شئ ، " المناطق الجينيّة غير المشفرة حُسبت نسبتها
98 % إلى 99 % من الجينوم البشرى وتتكون من إنترون وُجدت مع نصوص نسخ
البروتين ، و من مناطق بين-جينيّة بينهم " 25
أضف إلى تلك الأرقام (
هذه ) النتائج المدهشة جدا ، ( وهى ) أنّ عديد من التسلسلات الجينيّة (و)
التى لا تُتنج لا بروتين ولا آر إنّ إيه وُجدوا أن لهم وظيفة أيضا ( إنظر
نقاش البيكونز ) :

من كان يتوقع هذا ؟ __ هل هى بجانب الخلق ( أى الخلقيون) و أصحاب نظرية التصميم الذكى ؟
الخلقيون
وأصحاب نظريّة التصميم الذكى ادعوا لسنوات عديدة أنّ مفهوم " دآ إنّ إيه
الخردة" (( فضلا عن الهياكل الأثريّة ( أو اللاوظيفيّة )) لم يكن صحيحا
تماما .
لقد دعمت بنفسى هذه الفكرة لسنوات ( منذ 1997) .
حتى الآن ،
فقط الآن بدأ أخيرا تيار العلماء فى فهم الأخطاء المهمة فى إعتقادتهم التى
كانت مصدر إعتزازهم طويلا ، وذلك عندما تعلقت هذه الإعتقادات بفكرة الـ
دى إنّ إيه الخُردة الخاطئة _ هى فكرة كانت تقوم على الإقتراضات التطوريّة
المعقودة بحماس ، والتى أعمت تيار العلماء ومنعتهم من البحث_ لفترة طويلة
إلى حدّ ما _عن الكنوز الخفيّة لما يُسمى " دى إنّ إيه خُردة " .
عندما
بدأ العلماء فى إدراك أنّ نموذج الخلقيين لديه بالفعل قيمة علميّة تنبؤيّة
عالية ، وذلك عندما يتعلق الأمر بالفهم والتحقيق الدقيق ( فى ) العالم
المادى والكون .
إضافة إلى هذا ، وبالنظر فى نتائج " البيكنونز" Pyknonsالحديثة نسبيا (2006) بواسطة رجيوتوسس وآخرين . 24
البيكنونز
هم انماط فى تسلسل الـ دى إنّ إيه ذات أطوال مُتغيرة ( أى مختلفة ) ، و
التى لها نُسَخ مُتماثلة محفوظة و لها كثرة فوق ما يُتوقع بالصدفة ( أى
انها صدفة
إنهم أيضا لا يُنسخوا إلى آر إنّ إيه ( على عكس إم آر إنّ إيه المُشار إليه بالاعلى ) ، ولا يُترجموا إلى بروتين .
من
بين ملايين الأنماط المُكتشفة ، ريجيتوسس وآخرون وجدوا مجموعة فرعيّة ( أو
ثانويّة ) تتكون من 127.998 من الأنماط ، والذين أثطلق عليهم بيكينونز ،
والذين لديهم مثيلات ( اى شبههم ) _إضافيّة غير مُتداخلة _ فى المناطق
الغير مُترجمة و( مناطق) ترميز البروتين ، (الخاصة بـ ) 30.675 من النصوص ،
من عدد 20.059 من الجينوم البشرى .
البيكنوينز تُرتب إندماجيّا فى المناطق المُشفرة وغير المُترجمة لعديد من الجينات البشريّة ، حيث يُكونوا فُسيفساء .
المثيلات
المُتتابعة من البيوكينز فى هذه المناطق تُظهر ميلا( أو إنحيازا ) قويا
فى وضعيّة النسبيّة ، فهم يفضلون مسافات من 22 من النيكليوتيدات تقريبا.
البيكونيز أيضا شائعة فى الجينوم البشرى . إنّها تُشكل سُدس المناطق البين جينيّة والإنترونك ، أى ما مجموعه 127.998
بيكوينز
تُغطى 898.424.004 موقع من مواقع نيكليوتيد دآ إنّ إيه ، فى الخيوط
الاماميّة والعكسيّة ( المكونة للدى إن إيه ) للجينوم البشرى .
المُثير هنا ، بالبطبع ، أنّ البيكونزات مُرتبطة بعمليات بيلوجيّة مُحددة _ بمعنى ، أنّهم لهم وظيفة ..
المُقارنة
بين الجينات تكشف ان العديد من البيكونز لديهم مثيلات فى منطقة (NHGRI) من
جينات بعض الفقاريات الغيرفقاريات ، حيث هى مُمثله تمثيلا مُفرطا ( أى
مُتماثلة جدا ) فى العمليات البيولوجيّة ، كما فى الجينوم البشرى .
هذه "
االنتيجة الغير مُتوقعة " تُشير ، وفقا للمؤلف ، إلى وجود إرتباطات
وظيفيّة ممكنة فريدة من نوعها بين الأجزاء المُرمزة والغير مُرمزة من
الجينوم البشرى _ مثل إحتمال وجود إرتباط بين إسكات عملية بعد النسخ للجين
و تدخل آر إنّ إيه
البيكونزات البشريّة موجودة ايضا فى الجينومات الاُخرى ، حيث لهم إرتباط بعمليات بيولوجيّة مُماثلة .
والجدير
بالذكر ، أنّ أكثر من 600 مليون نيكليوتيد المُرتبطة بنُسخ البيكونزات
الغير جينيّة فى الجينوم البشرى ، مفقودين من جينوم القأر والجُرذ .
ومن
المثثير للإهتمام ، البيكونزات البشريّة لديهم عديد من المثيلات فى
المناطق البين جينيّة والإنترونك لجينوم دودة الفيلوجينتكال البعيدة وذبابة
الفاكهة ، التى تُغطى 1.6 مليون نيكلويتد فى كُلّ منهما تقريبا .
بالنظر
إلى التسلسلات الجينيّة التى تُرجمت او كُتبت أو ( حدث لها ) الإثنين ،
تبدوا أنّها محسوبة كـ " غالبية " الجينوم ، ويُظن أنّه مُفيدة وظيفيّا .
من المُثير للإهتمام أنّ انواع معيّنة من التسلسلات الجينيّة ، والتى هى لا تُرجمت ولا كُتبت ( أو نُسخت ) وُجد أيضا أنهم ذو وظيفة .
بالأخذ
القولين معا ، إنّه يبدوا أنّ الغالبيّة العُظمى من الجينوم -_ فى
الواقع _ وظيفيّة ، ولو على الأقل إلى درجة ما _ أكثر من 50% إن لم يكن
أكثر مثل 85 -90 % إن لم يكن أعلى ؟
مُفتاح الإختلافات بين القرد والإنسان .
لقد
أصبح أكثر وأكثر وضوحا أن الإختلافات الوظيفيّة الرئيسيّة بين الأشياء
الحيّة ، مثل الإنسان والقردة العُليا ، ليست موجودة بكثرة فى جينات ترميز
البروتين ، لكن فى المناطق الغير مُرمزة من ال دى إنّ إيه ، والتى كان
يُعتقد أنّها لا وظيفة لها " دى إن إيه خُردة " _ بقايا تطوريّة من أخطاء
الماضى ، والتى هى مُشتركة بين مُختلف المخلوقات .
هذه الفكرة بدأ
التخلص منها بمزيد ومزيد من الإكتشافات والتى بينت أن عديد من هذه المناطق
ليسوا فقط لهم وظيفة ، بل هم يحملوا الغالبيّة العُظمى من المعلومات
الجينيّة .
الـ " جينات " اللتى ذات مرة كان يُعتقد أنّها مهمة جدا
للوظيفة الجينيّة تحولوا إلى مُعادلين للّبنات الأساسيّة ذات المستوى
الأشد إنخفاضا فى الجينوم ، مثل الطوب السيارات (أو الموتور ) .
من
المُدهش ، أن المناطق الغير مُرمزة من الت دى إنّ إيه هى التى تتحكم فيما
سيحدث بهذه اللبنات _ والتى تُحدد أى نوع من " المنزل " يُبنى إذا جاز
التعبير .
المقالة التالية مُثيرة جدا للإهتمام فى هذا الصدد :- .
مراجع
ال إم آر إنّ آيه المُختلفة ، إضافة إلى الإختلافات فى مستويات تعبير ال
إم آر إنّ آيه المحفوظ ، يمكن أن يُساهم فى معرفة أسباب الإختلافات فى
التعبير الجينى المُلاحظة فى دماغ الإنسان والشامبنزى .
على الرغم من
الأهميّة الفسيولوجيّة التى ظهرت لل إم آر إنّ آيه فى مستوى منخفض أى لم
تظهر كلها ، إلّا أنه من المُغرى التكهن بأن مجموعمة من مثل هذا ال إم آر
إنّ آيه يُمكن أن تُساهم فى تنوع البرامج التطوريّة والعمليات الخلويّة .
كمثال ، ال إم آر إنّ آيه تبين ان لها علاقة حديثا فى التطورات المُتشابكة وفى تشكيل الذاكرة .
لأنّ الأنواع الخاصة من ال إم آر إنّ آيه الموصوفة هنا تم تعبيرها فى الدماغ ، والذى هو النسيج الأكثر تعقيدا فى جسم الإنسان ،
مع
ما يتراوح من 10.000 انواع مُختلفة من الخلايا ، هذه ال إم آر إنّ
آيهيُكمن أن يكون لها دور فى إنشاء أو المُحافظة ( أو إستمرارية ) على
التنوع الخلوى وبالتالى يمكن أن تُسهم فى الإختلافات بين دماغ الأنسان
والشامبنزى ... وظيفية ( أى وظيفيّا ) . 23
الجينات الزائفة وُجد أيضا أنّ لها وظائف مُماثلة للـ إم آر إنّ إيه .
"
نصوص الجينات الكاذبة المُعالجَة ربما تحتوى على مناطق بها مُتناظرات
مُتماثلة هامة ، والتى يمكن أن تُشير إلى دور تنظيمى للجينات الكاذبة
المُنسوخة من خلال ميكانيكيّة تُشبه الـ آر إنّ إيه " ( رابط :
http://www.clockss.org/clockss/Molecular_Interventions
)
هناك دراستان حديثتان أظهرتا أن مثل هذه الجينات الكاذبة المنسوخة تستطيع تنظيم النسخ لجينات ترميز البروتين المتناظرة .
نسخ
الجينات الزائفة فى ليمنيا إستجنالز ، والتى هى مُناظرة لجين مُصنّع
أُكسيد النيتريك ، تقلل مستويات التعبير للجين ، من خلال تكوين آر إن إيه
مزدوج ( او إزدواج ال آر إن إيه ) . هذا من المتوقع انه نشأ عن طريق تسلسل
عكسى مُكمّل موجود فى النهاية 5 من نص الجين الزائف . ( رابط :
)
وفى
مثال آخر ، فإنّ نسخ الـ makorin1-p1 TPΨg فى الفئران كان مطلوبا من اجل
إستقرار الـ إم آر إن إيه الذى هو من جين مُناظر makorin1.
هذا التنظيم أُستنج أنه نشأ من مجال فى مناطق الـ 5 لكل من الجينات والجينات الزائفة . ( رابط :
)
وفى الآونة الأخيرة ، ويل وآخرون إكتشفوا أن الجين الزائف FGFR-3 تم نسخه فى أنسجة الجنين فى إتجاه مُضاد ( أو مُضاد للإنتساخ
وهذا ما أوجب تقديم الإعتبارات القادمة :
كما
أن المناطق الدقيقة المماثلة بين FGFR-3 وجينها الزائف يمكن ان تصل إلى
طول 60 إن تى ، فإنّه من المُتوقع أنّ نصوص الـ FGFR-3 يمكن ان تلعب دورا
تنظيميا فى تعبير الـ FGFR-3 .
لو أن هذه النصوص المُضادة ( أو المضادة
للإنتساخ ) يمكن تهجينها إلى نصوص FGFR-3 ذات معنى داخل الخلايا ، هذا ربما
يؤدى إلى إمّا تدهور سريع أو تثبيط الترجمة .( رابط :
)
كما
أنّ ياو وآحرون توقعوا " المزيد من الدرسات عن الجينات الكاذبة المنسوخة (
او المُحولة ) سوف تُضيف لفهمنا لأدوراهم المُحتملة ، كما حدث مع جينات ال
آر إنّ إيه الغير مُرمّزة أو الأنواع الجديدة من العناصر الوظيفيّة " (
رابط :
)
إنّه
يبدوا أن كثيرا من الجينات الزائفة المنسوخة ربما تعمل كـ إم آر إن إيه
عملاقة ، لتنظيم وظيفة جينات ترميز البروتين والعناصر الجينيّة الأُخرى .
هناك
إختلافات مُثيرة أُخرى تشمل حقيقة أن أكثر من 6 % من الجينات بين الإنسان
والقردة العليا فريدة من نوعها سواء للإنسان أو للقردة العليا ، بمعنى ، لا
يُمكن مُشاركتها ( من نسب مشترك ) .
" نتائجنا تعنى أنّ البشر
والشامبنزى يختلفون على الأقل فى 6 % ((1.418 من 22.000 جين )) فى تكملة (
أو تمم ) الجينات ، والتى تقف فى تناقض صارخ مع ما يُستشهد به من إختلاف
1.5 %
بين التسلسلات الجينيّة المُتماثلة هذا " الباب الدوار "
الجينومى لربح وخسارة الجين يُمثل عددا كبيرا من الإختلافات الجينيّة التى
تفصل البشر من أقرب أقربائنا " 55sup>
وإذا كان أحد يعتقد ( أن وجود ) 6% اختلاف هو شئ مثير للاعجاب , فماذا عن وجود اختلاف فى أكثر من 30% ؟ مستحيل ؟ فكر مرة أخرى
وقد
أوضحت دراسة نُشرت فى أوائل عام 2010 أنه يظهر فقط هذا الفرق بين
الكروموسومات Y فى البشر والقردة , الكروموسوم Y عند الشمبانزى لم يُسلسل
كاملآ ولم يُعين مباشرة قبل إجراء الدراسة أبدآ . وأظهرت الدراسة وجود
اختلافات كثيرة مذهلة بين الهيكل والمحتوى الجينى للكروموسوم الإنسانى
وللشمبانزى , وحتى الجينات الفريدة نوعيآ بين النوعين . فيما يختص بالنظر
فى جينات معينة , يبدو أن هناك اختلاف هائل فى المحتوى الجينى للكروموسوم
Y- الخاص بالإنسان والشامبنزى – يصل إلى 53% . وبعبارة أخرى فإن الشمبانزى
يفتقر إلى ما يقرب من نصف الجينات الموجودة فى الكروموسوم Y الخاص بالإنسان
. ولأن الجينات تحدث فى العائلات أو الفئات المتشابهة فإن الباحثون قد
سعوا أيضآ لتحديد ما إذا كان هناك أى فرق فى فئات الجينات الفعلية . وقد
وجدوا 33% فرق مُروع . فالكروموسوم Y للإنسان يحتوى على نوع من الفئات
الجينية ثالثة إضافية _فئات مختلفة تمامآ _ مقارنة بالشمبانزى . 58 .
وينبغى
فى ظلّ الإفتراضات التطوريّة للتغيرات الجينيّة الطويلة والتدريجيّة ، ان
هيكل الكرموسوم Y ، التخطيطات ، الجينات ، والتسلسلات الأُخرى ينبغى أن
تكون مُتطابقة ( أو نفسها ) أكثر فى كلا النوعين ، وفى( أو بإعطاء ) 6
مليون سنة فقط أو قريبا من ذلك نظرا لأنّ من المُفترض أنّ الشامبزى
والإنسان ينحدرون من سلف مُشترك .
ولكن بدلا من ذلك فإنّ الإختلافات بين
كروموسوات واى ، والتى لُوحظت بواسطة آر . سكوت هولى ، وهو باحث فى علم
الوراثة فى معهد نيفيز فى مدينة كانساس ، وعلى الرغم من أنها لم تُشارك فى
البحث ، كما أخبر الإعلام المُصاحب ، " هذه النتيجة مُذهلة " . 59
لان
تقريبا كل جانب هيكلى من كروموسومات واى عن الإنسان والشامبنزى مختلف ،
فإنّه من الصعب التوصل إلى تقدير تشابه عام بينهما ، والذى لم يأخذ فى
الحُسبان غختلافات الحجم ، أو إختلافات الترتيب الهيكلى .
وقد تم
ذلك بأن تم الخلوص إلى أن فقط 70 % من تسلسل الشامبنزى ستكون مُتحاذية مع
تسلسل الإنسان _ دون اخذ الإختلافات فى المُحاذات فى الإعتبار .
وبعبارة
أُخرى ، فإنّ 70 % كانوا تقدير محفوظ ، خصوصا بإعتبار أنّ 50 % من الجينات
البشريّة مفقودة من الشامبنزى ، والمناطق المناطق التى بها لعض التشابه
كانت موجودة بأنماط مُختلفة تماما .
عندما تُؤخذ جميع جوانب عدم التشابه
( فئات السلسل ن الجينات ، أُسر الجينات ، موقع الجينات ) ، فسيكون آمنا
أن نقول أن التشابه هو أقل من 40 % حقيقة .
فى الحقيقة ، هذا الإختلاف
مُذهل جدا ، وهو ما جعل كُتاب مجلّة الطبيعة يصفوا التناقض مع النموذج
التطورى القياسى بطريقة مُثيرة إلى حدّ ما
FGFR-3 = *
خمسة وسبعون فى
المئة من ام اى ار ان ايه ( ميكرو آر إن آيه ) البشرية المعروفة والتى
أًستنسخت فى هذه الدراسة تم حفظها فى الفقاريات والثدييات , 14% حُفظت فى
اللافقاريات , 10% مختصة بالثدييات العليا ( الرئيسية ) و 1% مختصة
بالانسان .
ام اى ار ان ايه الجديدة لها توزيع حفظى مختلف : أكثر من نصف
الـ ام اى ار ان ايه البشرى تم حفظه فقط فى الرئسيات , حوالى 30% فى
الثدييات و9 % فى اللاثدييات الفقارية أو اللافقارية : 8% كانت مخصصة
للانسان . نحن نرى توزيع متشابه للـ ام ار ان ايه الخاص بالشمبانزى
References:
acq C, Miller JR, Brownlee GG. A
pseudogene structure in 5S DNA of Xenopus laevis,
Cell 12:109-120. 1977.
Gibson L. J., Pseudogenes and
Origins, Origins 21(2):91-108. 1994.
Menotti R.M., Starmer W.T., Sullivan D.T.,
Characterization of the structure and evolution of the Adh region of
Drosophila hydei,
Genetics 127:355-366. 1991.
Lalley P.A., Davisson M.T., Graves J.A.M., O’Brien S.J., Womack J.E.,
Roderick T.H., Creau-Goldberg N., Hillyard A.L., Doolittle D.P., Rogers
J.A., Report of the committee on
comparative mapping, Cytogenetics and Cell Genetics 51:503-532. 1989.
Long M., Langley C.H., Natural
selection and the orgin of jingwei, a chimeric processed functional gene in
Drosophila,
Science 260:91-95. 1993.
Jerlstrom, Pierre. 2000. Pseudogenes. Creation Ex Nihilo Technical
Journal 14 (no. 3):15.
Woodmorappe, John.2000. Are Pseudogenes 'Shared Mistakes' Between Primate
Genomes? Creation Ex Nihilo Technical Journal 14 (no. 3):58-71.
Abate, Tom. 2001. Genome Discovery Shocks Scientists. San Francisco
Chronicle (February 11).
Cantrell, Michael A. and others. 2001. An Ancient Retrovirus-like Element
Contains Hot Spots for SINE Insertion. Genetics 158:769-777.
Hillis, David M. 1999. SINEs of the perfect character. Proceedings of
the National Academy of Sciences 96:9979-9981.
Max, Edwards. Plagiarized Errors and Molecular Genetics.
Creation/Evolution (XIX, p.34) 1986-2003. (
http://www.talkorigins.org/faqs/molgen/
)
Lee, Jeannie T., Complicitiy of the gene and pseudogene, Nature
423:26-28. 2003
Hirotsun, Shinji et. al., An expressed pseudogene regulates the
messenger-RNA stability of its homologous coding gene, Nature
423:91-96. 2003
Makalowski, Wojciech.
2003. Not Junk After All, Science 300:1246-1247
Wyatt Gibbs, The Unseen Genome: Gems among the Junk, Scientific
American, November 2003, pp 45-53 (
Link
)
ENCORE Project Consortium et al., Identification and analysis of functional
elements in 1% of the human genome by the ENCODE pilot project,
Nature
447, 799-816 (14 June 2007); Richard Ingham, Landmark study prompts rethink
of genetic code, Yahoo News, accessed June 15, 2007 (Link1,
Link2)
Nishikimi, M. and Yagi, K. (1991) Molecular basis for the deficiency in
humans of gulonolactone oxidase, a key enzyme for ascorbic acid
biosynthesis. Am. J. Clin. Nutr. 54(6 Suppl):1203S-1208S.
Nishikimi, M., Fukuyama, R., Minoshima, S., Shimizu, N. and Yagi. K. (1994)
Cloning and chromosomal mapping of the human nonfunctional gene for
L-gulono-gamma-lactone oxidase, the enzyme for L-ascorbic acid biosynthesis
missing in man. J. Biol. Chem. 269:13685-13688.
Ohta, Y. and Nishikimi, M. (1999) Random nucleotide substitutions in primate
nonfunctional gene for L-gulono-gamma-lactone oxidase, the missing enzyme in
L-ascorbic acid biosynthesis. Biochim. Biophys. Acta. 1472:408-411.
Inai, Y., Ohta. Y., and Nishikimi, M. (2003) The whole structure of the
human nonfunctional L-gulono-gamma-lactone oxidase gene--the gene
responsible for scurvy--and the evolution of repetitive sequences thereon.
J Nutr Sci Vitaminol (Tokyo) 49:315-319.
Peter Borger, Shared mutations: Common descent or common mechanism?, The
Independent Research Institute on Origins, Accessed 8/10/07 (
Link
)
Eugene Berezikov, Fritz Thuemmler, Linda W van Laake, Ivanela Kondova,
Ronald Bontrop4, Edwin Cuppen & Ronald H A Plasterk, "Diversity of microRNAs
in human and chimpanzee brain", Nature Genetics, Vol 38 | Number 12 |
December 2006 pp. 1375-1377. (
Link
)
Isidore Rigoutsos, Tien
Huynh,
Kevin Miranda, Aristotelis Tsirigos, Alice
McHardy, and Daniel Platt, Short blocks from the
noncoding parts of the human genome have instances within nearly all known
genes and relate to biological processes, PNAS | April 25, 2006 | vol. 103 |
no. 17 | 6605-6610 (
Link
)
Jill Cheng,
Philipp Kapranov, Jorg Drenkow,
Sujit Dike, Shane Brubaker, Sandeep Patel,
Jeffrey Long, David Stern, Hari Tammana,
Gregg Helt, Victor Sementchenko, Antonio
Piccolboni, Stefan Bekiranov, Dione K. Bailey,
Madhavan Ganesh, Srinka Ghosh, Ian Bell,1
Daniela S. Gerhard,
Thomas R. Gingeras,
Transcriptional
Maps of 10 Human Chromosomes at 5-Nucleotide Resolution, Science 20
May 2005: Vol. 308. no. 5725, pp. 1149 - 1154 (
Link
)
Richard Twyman, Small RNA: BIG NEWS, The Human Genome, January 2005 (
Link
)
J. J. Bull, M. R. Badgett, H. A. Wichman, J. P. Huelsenbeck, D. M. Hillis,
A. Gulati, C. Ho, and I. J. Molineux, Exceptional Convergent Evolution in a
Virus, Genetics, 1997
December; 147(4):
1497–1507. (
Link
)
Jose´ M. Cuevas, Santiago F. Elena and Andre´s Moya, Molecular Basis of
Adaptive Convergence in Experimental Populations of RNA Viruses, Genetics,
October 2002, 162: 533–542 (
Link
)
H A Wichman, L A Scott, C D Yarber, and J J Bull, Experimental evolution
recapitulates natural evolution,
Philos Trans R Soc Lond B Biol Sci.
2000 November 29; 355(1403):
1677–1684. (
Link
)
Troy E. Wood, John M. Burke and Loren H. Rieseberg, Parallel genotypic
adaptation: when evolution repeats itself, Genetica, February 2005, Volume
123, Numbers 1-2, pp. 157-170 (
Link
)
F. Flam, Hints of a language in junk DNA, Science 266:1320, 1994.
Haussler and Gill Bejerano, Junk DNA, May 6,
2004 online version of
Science.
(
Link
)
Stanford University Medical Center (2007, April 24). 'Junk' DNA Now Looks
Like Powerful Regulator, Scientists Find. ScienceDaily. (
Link
)
Nóbrega MA, Zhu Y, Plajzer-Frick I, Afzal V, Rubin EM., Megabase deletions
of gene deserts result in viable mice, Nature.
2004 Oct 21;431(7011):988-93. (
Link
)
Katherine Dunlap, et at., Endogenous
retroviruses regulate periimplantation placental growth and differentiation,
PNAS
September 26, 2006 vol. 103
no. 39 14390-14395 (
Link
)
Lori F. Maxfield, Camilla D. Franze, John M. Coffin, Relationship between
retroviral DNA-integration-site selection and host cell transcription,
PNAS
February 1, 2005 vol. 102
no. 5 1436-1441 ( Link
)
Sverdlov, E. D. (2000) "Retroviruses and primate evolution." BioEssays 22:
161-171. (
Link
)
Luis P. Villarreal, The Viruses That Make Us: A Role for Endogenous
Retroviruses in the Evolution of Placental Species, UCI Online Faculty
Publication (
Link: last accesses 3/10/09).
Polavarapu N, Bowen NJ, McDonald JF.
Identification, characterization and comparative genomics of chimpanzee
endogenous retroviruses. Genome Biol. 2006;7(6):R51.
Barbulescu M, Turner G, Su M, Kim R, Jensen-Seaman MI, Deinard AS, Kidd KK,
Lenz J., A HERV-K provirus in chimpanzees, bonobos and gorillas, but not
humans. Curr Biol. 2001 May 15;11(10):779-83.
Patric Jern, Göran O. Sperber, and Jonas Blomberg, Divergent
Patterns of Recent Retroviral Integrations in the Human and Chimpanzee
Genomes: Probable Transmissions between Other Primates and Chimpanzees,
Journal of Virology, February 2006, p. 1367-1375, Vol. 80, No. 3
(
Link
- last accessed 3/11/09 )
Chris T Yohn,#1 Zhaoshi Jiang,#2 Sean D McGrath,2
Karen E Hayden,1 Philipp Khaitovich,3 Matthew E
Johnson,1,2 Marla Y Eichler,2 John D McPherson,4
Shaying Zhao,5 Svante Pääbo,3 and Evan E Eichler,
Lineage-Specific Expansions of Retroviral Insertions within the Genomes of
African Great Apes but Not Humans and Orangutans,
PLoS Biol. 2005 April;
3(4):
e110.
Sverdlov, E. D. (2000) “Retroviruses and primate evolution.” BioEssays
22:161-171. ( Link
)
Nicholas H. Putnam,1
Mansi Srivastava,2
Uffe Hellsten,1 Bill Dirks,2
Jarrod Chapman,1 Asaf Salamov,1
Astrid Terry,1 Harris Shapiro,1
Erika Lindquist,1 Vladimir V. Kapitonov,3
Jerzy Jurka,3 Grigory Genikhovich,4
Igor V. Grigoriev,1 Susan M. Lucas,1
Robert E. Steele,5 John R. Finnerty,6
Ulrich Technau,4 Mark Q. Martindale,7
Daniel S. Rokhsar,
Sea Anemone Genome Reveals Ancestral
Eumetazoan Gene Repertoire and Genomic Organization, Science 6 July
2007: Vol. 317. no. 5834, pp. 86 - 94 (
Link
) (see also the following review of this article:
Link
)
David Posada and Keith A. Crandall, Simple (Wrong)
Models for Complex Trees: A Case from Retroviridae, Molecular Biology
and Evolution
18:271-275 (2001) (
Link
)
Andrew B. Conley, Jittima Piriyapongsa and I. King Jordan, "Retroviral
promoters in the human genome," Bioinformatics, Vol. 24(14):1563–1567
(2008) ( Link
)
Richard Sternberg, "On the Roles of Repetitive DNA Elements in the Context
of a Unified Genomic–Epigenetic System," Annals of the New York Academy
of Sciences, Vol. 981: 154–88 (2002)
University of California, Santa Cruz, Ancient retroviruses spurred evolution
of gene regulatory networks in humans and other primates, Online News
Article, Physorg.com, November 14th, 2007 (
Link
- last accessed 3/13/09 )
Alex D Greenwood, Claudia C Englbrecht, and Ross DE MacPhee,
Characterization of an endogenous retrovirus class in elephants and their
relatives, BMC Evol Biol.
2004; 4:
38. (
Link
)
Manuela Mura, et al., Late viral interference induced by
transdominant Gag of an endogenous retrovirus,
PNAS,
July 27, 2004 vol. 101 no. 30 (
Link
)
Norbert Bannert and Reinhard Kurth, Retroelements and the human genome: New
perspectives on an old relation,
PNAS
October 5, 2004, vol. 101 no. suppl 2 (
Link
)
Jennifer F. Hughes and John M. Coffin, Human endogenous retrovirus K
solo-LTR formation and insertional polymorphisms: Implications for human and
viral evolution,
PNAS
February 10, 2004 vol. 101 no. 6 (
Link
)
IJdo JW, Baldini A, Ward DC, Reeders ST, Wells RA, Origin of human
chromosome 2: an ancestral telomere-telomere fusion. Proc Natl Acad Sci U S
A 1991 Oct 15;88(20):9051-5; (
Link
)
Douglas L. T. Rohde, Steve Olson & Joseph T. Chang, Modelling the recent
common ancestry of all living humans, Nature 431, 562-566 (30
September 2004) | doi:10.1038/nature02842;
Received 30 December 2003; Accepted 14 July 2004
Demuth JP, Bie TD, Stajich JE, Cristianini N, Hahn MW (2006) The Evolution
of Mammalian Gene Families. PLoS ONE 1(1): e85.
doi:10.1371/journal.pone.0000085 (
Link
)
Li et al. Extensive, Recent Intron Gains in Daphnia Populations. Science,
2009; 326 (5957): 1260 DOI:
10.1126/science.1179302
| See also: Indiana University (2009, December 14). Introns --
nonsense DNA -- may be more important to evolution of genomes than thought.
ScienceDaily. Retrieved December 15, 2009,
(
Link
).
Erika Check Hayden, Human genome at ten: Life is complicated,
Nature
464, 664-667, Published online
31 March 2010 (
Link
)
Hughes, J.F. et al. 2010. Chimpanzee and human Y chromosomes are remarkably
divergent in structure gene content. Nature. 463 (7280): 536-539. (
Link
).
Borenstein, S. Men More Evolved? Y Chromosome Study Stirs Debate.
Associated Press, January 13, 2010. (
Link
)
Richard Sternberg, Guy Walks Into a Bar and Thinks He's a Chimpanzee: The
Unbearable Lightness of Chimp-Human Genome Similarity, Evolution News and
Views, May 14, 2009 (
Link
)
Wellcome Trust. "Scientists map hotspots for genetic exchange in
chimpanzees." ScienceDaily,
15 Mar. 2012. Web. 9 Apr. 2012. (
Link ); see also: A.
Auton, A. Fledel-Alon, S. Pfeifer, O. Venn, L. Segurel, T. Street, E. M.
Leffler, R. Bowden, I. Aneas, J. Broxholme, P. Humburg, Z. Iqbal, G. Lunter,
J. Maller, R. D. Hernandez, C. Melton, A. Venkat, M. A. Nobrega, R. Bontrop,
S. Myers, P. Donnelly, M. Przeworski, G. McVean. A Fine-Scale Chimpanzee
Genetic Map from Population Sequencing.
Science, 2012; DOI:
10.1126/science.1216872
The Chimpanzee Sequencing and Analysis Consortium,
Initial sequence of the chimpanzee genome and comparison with the human
genome, Nature 437,
69-87 (1 September 2005) (Link)
Andrew B. Conley, Jittima
Piriyapongsa and I. King Jordan, "Retroviral
promoters in the human genome,"
Bioinformatics, Vol.
24(14):1563--1567 (2008).


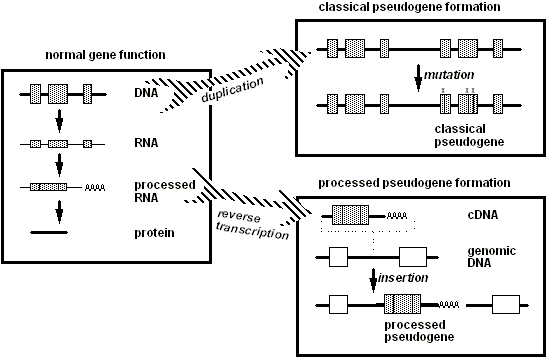


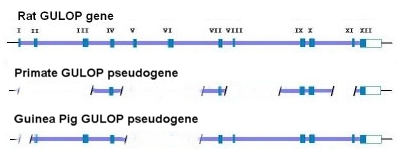

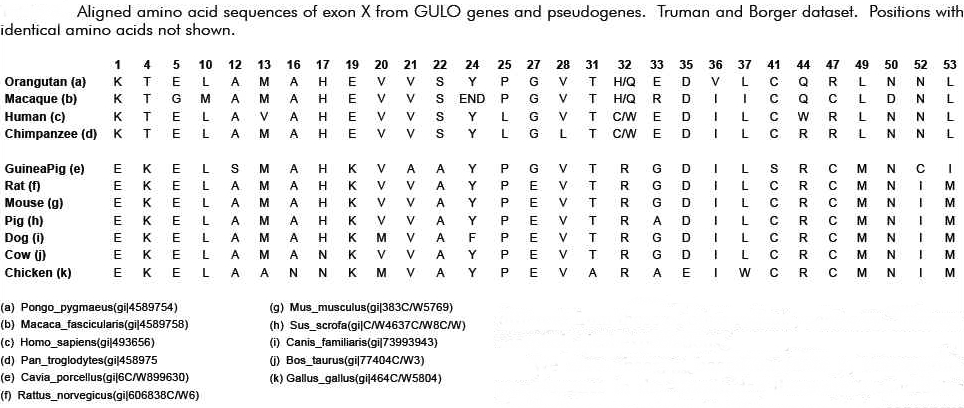

ليست هناك تعليقات:
إرسال تعليق